






關(guān)聯(lián)CD4 +/CD8+ T細(xì)胞新抗原疫苗克服免疫檢查點(diǎn)阻斷劑抗藥性,實(shí)現(xiàn)腫瘤消退

免疫檢查點(diǎn)阻斷療法(ICB)的治療效果目前僅限于被認(rèn)為具有足夠腫瘤突變負(fù)荷(TMB)的癌癥亞群,這些亞群允許自體T細(xì)胞自發(fā)識(shí)別新抗原(NeoAg)。本研究探討了是否可以通過(guò)聯(lián)合免疫療法改善侵襲性低TMB鱗狀細(xì)胞腫瘤對(duì)ICB的反應(yīng),該療法使用功能明確的NeoAg作為內(nèi)源性CD4+和CD8+T細(xì)胞的靶點(diǎn)。單獨(dú)接種CD4+或CD8+ NeoAg不能提供預(yù)防性或治療性免疫力,但含有這兩個(gè)亞群都能識(shí)別的NeoAg的疫苗能克服ICB抗性,并能根除包含PD-L1+ 腫瘤誘導(dǎo)癌干細(xì)胞(tCSC)亞群的已形成的大型腫瘤,前提是相關(guān)表位存在物理聯(lián)系。機(jī)制上,治療性 CD4+/CD8+ T 細(xì)胞 NeoAg 疫苗接種可改變腫瘤微環(huán)境 (TME),通過(guò)結(jié)合 ICB 介導(dǎo)的分子間表位擴(kuò)散,使處于祖細(xì)胞和中間衰竭狀態(tài)的 NeoAg 特異性 CD8+ T 細(xì)胞數(shù)量增加。該研究于2023年9月發(fā)表在《The Journal of Clinical Investigation》,IF:15.9。
技術(shù)路線:
主要研究結(jié)果:
1. 基于內(nèi)源性CD4+和CD8+T細(xì)胞反應(yīng)性的新抗原功能鑒定
用 1×107個(gè)照射過(guò)的SCC VII細(xì)胞(單獨(dú)或輔以 50 μg polyI:C)對(duì)C3H/HeJ 小鼠進(jìn)行皮下注射免疫。免疫小鼠在14天后再接種 5×105個(gè)轉(zhuǎn)導(dǎo)表達(dá)熒光素酶和綠色熒光蛋白(SCC VII-Luc/GFP)的 SCC VII 活細(xì)胞,以實(shí)現(xiàn)生物發(fā)光(BLI)追蹤。雖然僅用輻照過(guò)的 SCC VII 進(jìn)行全細(xì)胞免疫接種并不能保護(hù)小鼠在受到挑戰(zhàn)后免受腫瘤生長(zhǎng)的影響--根據(jù)這一經(jīng)典定義,SCC VII 是一種免疫原性很差的腫瘤--但通過(guò)多聚I:C的編碼遞送可以實(shí)現(xiàn)預(yù)防(圖1A和B)。因此,SCC VII 含有能夠產(chǎn)生保護(hù)性免疫的抗原。這取決于CD4+和CD8+ T 細(xì)胞,因?yàn)樵诮臃N疫苗之前(圖1C)或之后(圖1D),任何一個(gè)亞群的耗竭都會(huì)導(dǎo)致腫瘤在隨后的挑戰(zhàn)中生長(zhǎng)。值得注意的是,與對(duì)照組相比,在接種前就被耗盡CD4+ T細(xì)胞的小鼠的腫瘤生長(zhǎng)率降低,這表明在疫苗誘導(dǎo)反應(yīng)的起始階段和接種后維持其療效都需要這種亞群。
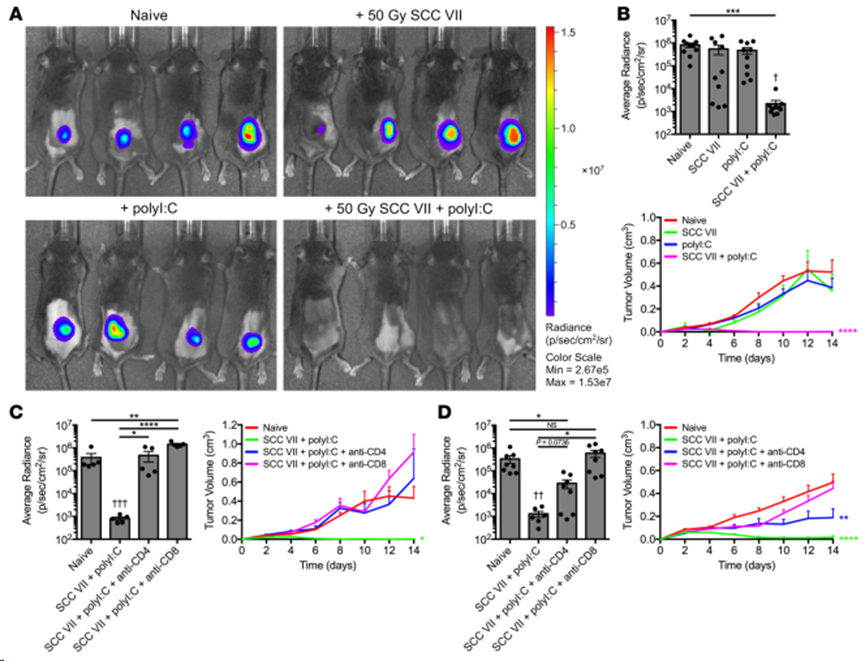
圖1. SCC VII和polyI:C聯(lián)合免疫對(duì)肝臟腫瘤的侵襲具有保護(hù)作用
隨后,對(duì)產(chǎn)生強(qiáng)烈 IFN-γ 反應(yīng)的肽池進(jìn)行去卷積,以檢測(cè)所針對(duì)的特定突變肽。分析結(jié)果顯示,SCC VII 的天然免疫反應(yīng)識(shí)別出的突變基因包括 Pik3ca(Mut_44)、Cltc(Mut_48)、Ctnnd1(Mut_61)和 Otud5(Mut_65 和 Mut_67)(圖2A和B)。鑒定體細(xì)胞變異、選擇候選突變進(jìn)行功能測(cè)試以及對(duì) NeoAg 進(jìn)行功能驗(yàn)證所涉及的步驟用Circos圖表示(圖2C)。
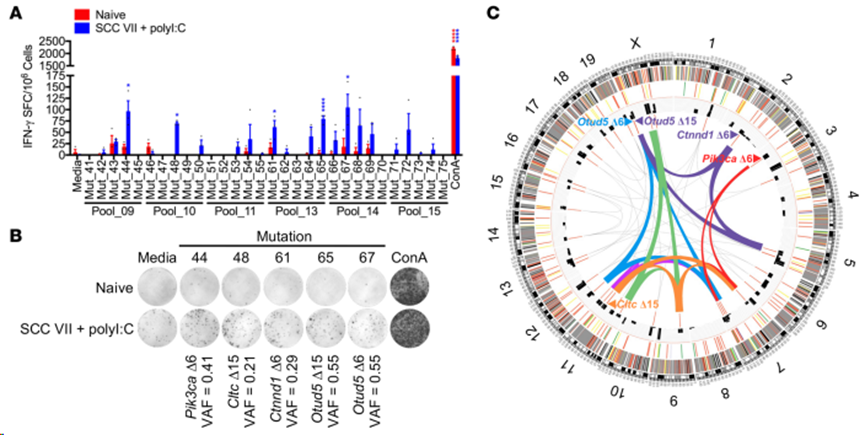
圖2. SCC VII和polyI:C聯(lián)合免疫后基于功能的新抗原鑒定
在檢測(cè)各NeoAg肽對(duì)觀察到的免疫的貢獻(xiàn)時(shí),只有Mut_48 (CltcΔ15)顯示出對(duì)攻擊產(chǎn)生保護(hù)作用(圖3A),而WT肽(WT_48)無(wú)保護(hù)作用(圖3B)。這些數(shù)據(jù)表明T細(xì)胞對(duì)Mut_48的反應(yīng)介導(dǎo)了預(yù)防性肽疫苗接種后的保護(hù)性免疫。
評(píng)估分離自SCCⅶ腫瘤細(xì)胞免疫小鼠的CD4+和CD8+ T細(xì)胞的反應(yīng)性,確定參與天然NeoAg特異性免疫應(yīng)答的T細(xì)胞亞群。發(fā)現(xiàn),Mut_48被CD4+和CD8+ T細(xì)胞識(shí)別,而Mut_44、Mut_61、Mut_65和Mut_67只能被CD4+ T細(xì)胞識(shí)別。此外,分離出的CD8+ T細(xì)胞識(shí)別Mut_72和Mut_73,這兩種肽在Slc26a11基因中含有相同的錯(cuò)義突變(圖3D)。然而,在初次/加強(qiáng)接種后,Mut_72和Mut_73均無(wú)法在體內(nèi)產(chǎn)生針對(duì)SCCⅶ的保護(hù)性免疫(圖3C)。這些結(jié)果表明,只有能被CD4+和CD8+ T細(xì)胞識(shí)別的Mut_48能夠通過(guò)多肽疫苗誘導(dǎo)有效的預(yù)防性免疫
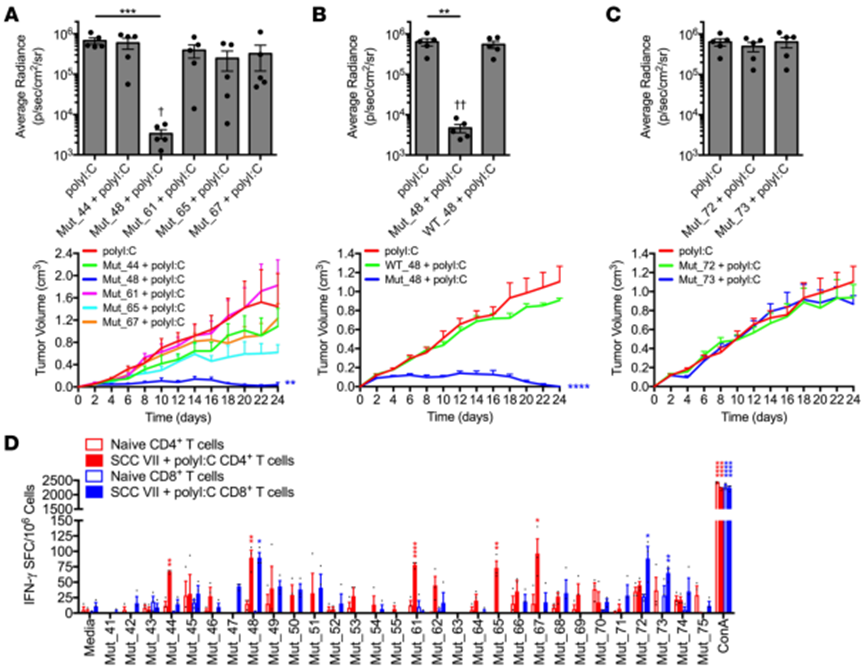
圖3. CD4+和CD8+ T細(xì)胞對(duì)SCC vii衍生的新抗原反應(yīng)的去卷積
為確定每個(gè)亞群識(shí)別的表位是相同的還是不同的,使用 IFN-γ ELISPOT 定量對(duì)一組含有Mut_48 H129Q突變的10-mer和15-mer肽(命名為Mut_48.1-Mut_48.10)的相對(duì)反應(yīng)量(圖4A)。從 SCC VII/polyI:C 免疫小鼠體內(nèi)分離出的 CD8+ T 細(xì)胞在識(shí)別 Mut_48.10 10-mer 后產(chǎn)生的 IFN-γ 量最大,抗體阻斷表明它是由 H-2Kk 呈遞的(圖4B)。共純化的CD4+ T 細(xì)胞對(duì)通過(guò) I-Ak 呈遞的 Mut_48.5 15-mer(圖4C)表現(xiàn)出最大的反應(yīng)性,而對(duì)任何設(shè)計(jì)的 10-mer都沒(méi)有預(yù)期的反應(yīng)(數(shù)據(jù)未顯示)。此外,使用免疫表位數(shù)據(jù)庫(kù)和分析資源(IEDB)NetMHCpan(v4.0)方法對(duì) Mut_48衍生的 10-mer與H-2Kk的結(jié)合進(jìn)行了硅預(yù)測(cè),估計(jì)大多數(shù)肽的親和力較差,Mut_48.10 的最佳親和力為 4988.7 nM IC50。值得注意的是,盡管預(yù)測(cè)的 H-2Kk 結(jié)合 IC50 值高于大多數(shù)篩選方案使用的 500 nM 臨界值(圖4D),但Mut_48.7-48.10 系列中包含的 10-mer肽仍能誘導(dǎo) IFN-γ 生成。這些結(jié)果進(jìn)一步證實(shí),Mut_48 在較長(zhǎng)的CD4+ T 細(xì)胞表位Mut_48.5 中包含一個(gè) CD8+ T 細(xì)胞最小表位Mut_48.10,從而賦予這兩個(gè)T細(xì)胞亞群產(chǎn)生 IFN-γ 的能力(圖4E)。對(duì)刺激IFN-γ 的Mut_72和Mut_73的H-2Kk 結(jié)合預(yù)測(cè)進(jìn)行擴(kuò)展分析后,發(fā)現(xiàn)這兩種肽的IC50親和力均為250.9 nM,這表明IEDB NetMHCpan(v4.0)工具算法在過(guò)濾 CD8+ T 細(xì)胞體外免疫活性方面的效率約為 66%(圖4F)。因此,本研究利用無(wú)偏見(jiàn)的功能性方法鑒定 NeoAg 使其能夠在同一檢測(cè)系統(tǒng)中更有效地探測(cè)CD4+和CD8+ T細(xì)胞表位。
免疫研究表明,含有CD4+和CD8+ T細(xì)胞識(shí)別的最小表位的Mut_48.5 15-聚合體對(duì)體內(nèi)活 SCC VII 細(xì)胞挑戰(zhàn)的保護(hù)程度與 Mut_48 20-聚合體相當(dāng)。此外,CD4+ T 細(xì)胞表位對(duì)于觀察到的保護(hù)作用是完全必要的,因?yàn)橛脙H含有 CD8+ T 細(xì)胞表位的截短 Mut_48.10 10-mer 免疫小鼠有部分保護(hù)作用(圖 4G)。
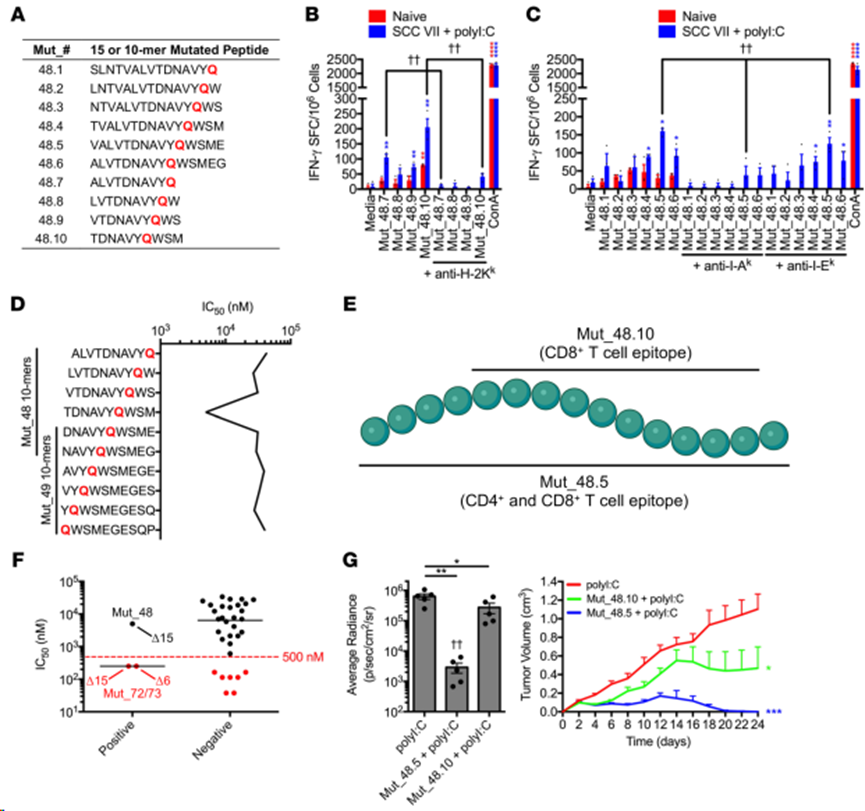
圖4. CD4+和CD8+ T細(xì)胞疫苗衍生表位的MHC限制和功能相互作用
2. CD4+T細(xì)胞的腫瘤特異性和對(duì)CD8+T細(xì)胞的幫助
通過(guò)三重丙氨酸重復(fù)(-AAA-)共價(jià)連接 CD4+ 和 CD8+ T 細(xì)胞抗原,與由無(wú)系肽組成的疫苗接種相比,Mut_48 對(duì) SCC VII 挑戰(zhàn)的保護(hù)效果更佳(圖 5)。因此,Mut_48 的功效與 CD4+ T 細(xì)胞輔助抗原以及腫瘤特異性 CTL NeoAg 的呈現(xiàn)有關(guān),很可能是由相同的APC呈現(xiàn)的。此外,這些結(jié)果表明,CD4+ T 細(xì)胞需要腫瘤特異性的其他效應(yīng)功能在該模型中是可有可無(wú)的。
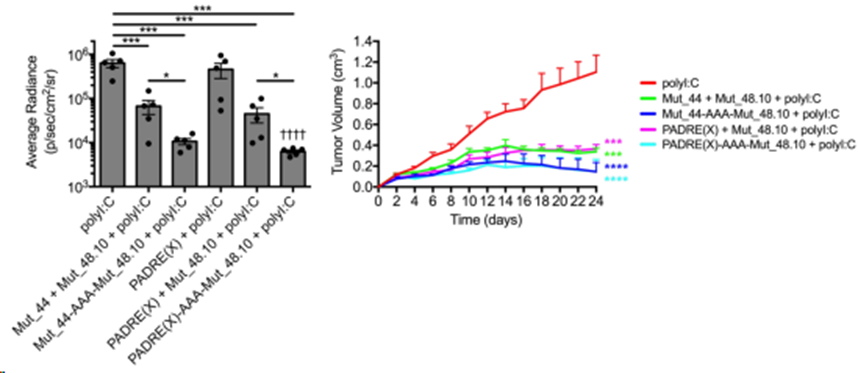
圖5. 栓系CD4+ T細(xì)胞輔助性和最小CD8+ T細(xì)胞表位疫苗盡可能抑制SCCⅶ腫瘤生長(zhǎng)
3. 結(jié)合新抗原疫苗接種可克服免疫檢查點(diǎn)阻斷的治療耐藥性
有研究表明,ICB 單藥治療可擴(kuò)大內(nèi)源性 NeoAg 特異性 T 細(xì)胞反應(yīng),并在與基于預(yù)測(cè)的疫苗聯(lián)合使用時(shí)產(chǎn)生新的 NeoAg 反應(yīng),從而提高患者的無(wú)進(jìn)展生存期。評(píng)估了基于已驗(yàn)證靶點(diǎn)的 NeoAg 疫苗能否與這一策略合理結(jié)合。首先使用上述原代/加強(qiáng)方案,通過(guò)免疫集合 NeoAg 疫苗(靶向 Pik3ca、Cltc、Ctnnd1 和 Otud5 突變)誘導(dǎo) NeoAg 特異性 T 細(xì)胞應(yīng)答。活體 SCC VII-Luc/GFP 腫瘤挑戰(zhàn)三天后,小鼠通過(guò)靜脈注射 PD-1 或 CTLA-4 阻斷抗體,并測(cè)量其對(duì)腫瘤生長(zhǎng)的影響。在這兩種情況下,與單獨(dú)使用 ICB 相比,ICB 都能明顯加快 NeoAg 疫苗對(duì)生長(zhǎng)中的 SCC VII 腫瘤產(chǎn)生治療性免疫的能力。將 ICB 與多肽疫苗結(jié)合使用可防止約 50% 的單獨(dú)接種 NeoAg 的C3H/HeJ小鼠晚期(超過(guò)第 24 天)SCC VII 腫瘤復(fù)發(fā)(圖 6A 和 B)。抗PD-1和NeoAg聯(lián)合疫苗接種明顯加速了可觸及腫瘤的消除,在第14天活動(dòng)性排斥的早期動(dòng)力學(xué)階段協(xié)同效應(yīng)明顯(圖6A)。此外,通過(guò)IFN-γ ELISPOT檢測(cè)NeoAg疫苗接種后第42天的記憶T細(xì)胞反應(yīng),發(fā)現(xiàn)PD-1阻斷增加Mut_48特異性T細(xì)胞反應(yīng)的幅度,并顯示分子間表位擴(kuò)散到Mut_72和Mut_73的證據(jù)(圖6C和D),這些靶點(diǎn)沒(méi)有包括在多肽疫苗接種中,但以前曾觀察到它們?cè)谂cCD4+ T細(xì)胞物理分離后引起CD8+ T細(xì)胞反應(yīng)(圖3D)。相反,抗 CTLA-4 處理既沒(méi)有顯示協(xié)同作用,也沒(méi)有顯著影響 Mut_48 特異性 T 細(xì)胞的絕對(duì)數(shù)量。此外,抗 CTLA-4 還減少表位向其他特異性的擴(kuò)散(圖 6B)。
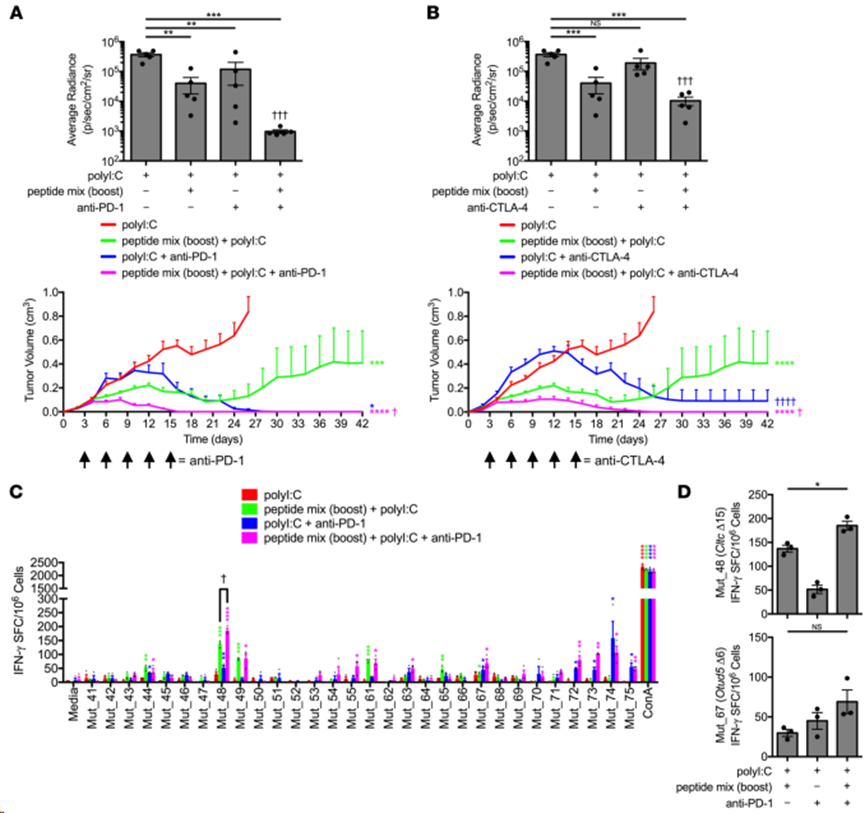
圖6. 抗PD-1檢查點(diǎn)阻斷增加CltcΔ15-specific記憶頻率,并促進(jìn)分子間優(yōu)勢(shì)表位的擴(kuò)散
SCC VII-Luc/GFP腫瘤在小鼠群中生長(zhǎng),使其體積達(dá)到約300至400立方毫米,然后在第10天和第24天用對(duì)側(cè)靜脈注射Mut_48+polyI:C混合物和/或靜脈注射抗PD-1治療2個(gè)周期。單獨(dú)使用Mut_48疫苗并不能帶來(lái)治療效果,而抗PD-1則表現(xiàn)出不同程度的原發(fā)性和繼發(fā)性耐藥性,有時(shí)只能導(dǎo)致最初的腫瘤控制,但隨后就會(huì)喪失。與此相反,將 PD-1 阻斷與 Mut_48 NeoAg 疫苗結(jié)合使用,可以完全、持久(超過(guò)90天)地根除已形成的大腫瘤(圖7A-D)。體外再刺激脾臟和腫瘤引流Ig LN的淋巴細(xì)胞發(fā)現(xiàn),聯(lián)合使用NeoAg和抗PD-1可協(xié)同增強(qiáng)記憶期Mut_48特異性T細(xì)胞反應(yīng)(圖7E)。此外,在第一輪免疫療法后的第 17 天效應(yīng)期分離的腫瘤浸潤(rùn)淋巴細(xì)胞(TIL)組分中,NeoAg 和抗 PD-1 聯(lián)合給藥顯著增加總 CD8+ T 細(xì)胞的數(shù)量,而常規(guī)(Tconv)和調(diào)節(jié)性(Treg)CD4+ T 細(xì)胞的數(shù)量保持不變(圖7F)。Mut_48 和抗 PD-1 治療對(duì)腫瘤的排斥也伴隨著對(duì) SCC VII 向區(qū)域 LN 轉(zhuǎn)移的抑制(圖7G)。這些數(shù)據(jù)表明,功能性 NeoAg 介導(dǎo)的腫瘤排斥和區(qū)域轉(zhuǎn)移預(yù)防在與 ICB 結(jié)合使用后具有最佳治療效果。
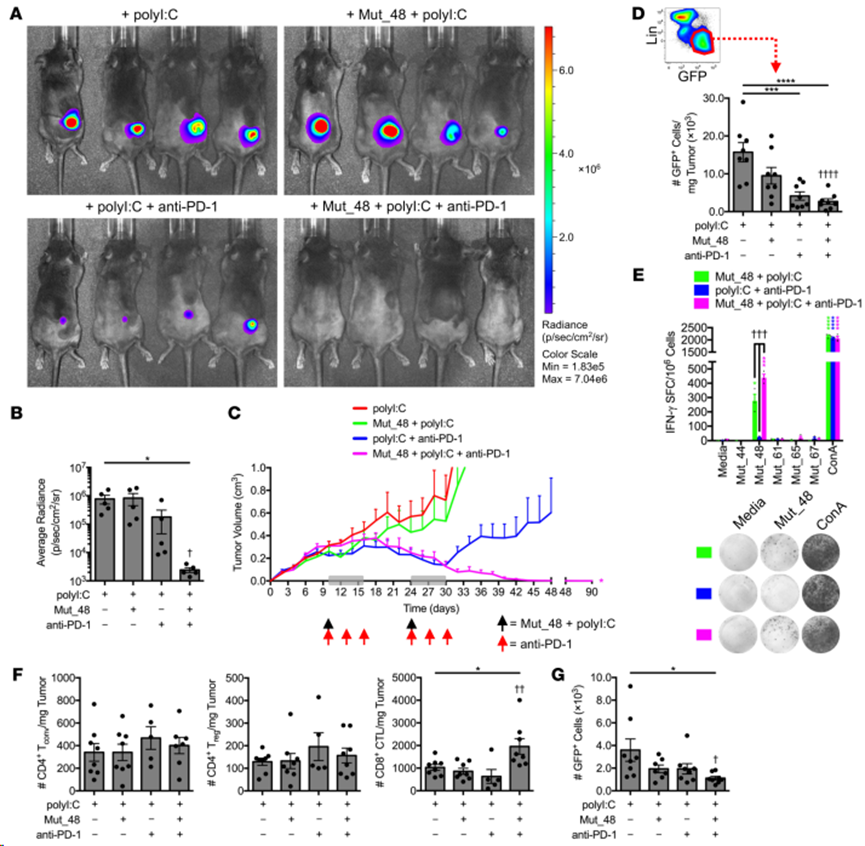
圖7. 抗PD-1和CltcΔ15的延遲聯(lián)合治療可促進(jìn)已建立的SCCⅶ腫瘤的清除
4. 新抗原疫苗接種增加干細(xì)胞樣和中間耗竭的CD8+T細(xì)胞的存在
在腫瘤接種后第 10 天,用 Mut_48 + polyI:C混合物和/或i.p.抗PD-1對(duì) SCC VII 腫瘤小鼠進(jìn)行s.c.處理,在第 17 天效應(yīng)期從 TIL、脾細(xì)胞和腫瘤排出的 Ig LN 分餾物中純化 CD45+ 細(xì)胞,并進(jìn)行 FACS 處理。將來(lái)自所有器官的入選 T 細(xì)胞合并在一起,并使用 21 種已知的表型特征將其投射到UMAP空間,以定義naive CD4+/CD8+ T 細(xì)胞(Tn)、CD4+/CD8+ Teff/mem、CD4+ Treg、CD8+ Tprec/prog、CD8+ Tex-int 和 CD8+ Tex-term 亞群。總CD4+和CD8+T 細(xì)胞得到清晰的分離(圖 8A)。來(lái)自 TIL、脾臟和 LN 的 CD8+ T 細(xì)胞亞群似乎完全不同,而來(lái)自TIL和LN的CD4+ T細(xì)胞似乎占據(jù)與脾臟類似的獨(dú)立空間(圖8B上)。當(dāng)單獨(dú)觀察TIL定位與治療的關(guān)系時(shí),可以觀察到Mut_48疫苗接種與特定CD4+和CD8+T細(xì)胞亞群消失之間的密切聯(lián)系,而相對(duì)于單獨(dú)的polyI:C治療,抗PD-1治療要么引起更微妙的變化,要么似乎擴(kuò)大先前存在的亞群(圖 8B下)。
為更詳細(xì)地了解治療是如何影響 T 細(xì)胞分化的,確定24個(gè)元集群,它們捕捉到在整個(gè) UMAP 區(qū)域觀察到的標(biāo)記物特征的粒度(圖8C和D)。該圖譜顯示,各器官的治療捕獲了CD8+Tex、Teff 和Tmem系,其中門控CD8+ T 細(xì)胞的偽時(shí)間軌跡分析顯示CD62L(Tn和Tmem)、TCF-1/SLAMF6(Tprec/prog 和 Tmem)、GzmB/CX3CR1/CD44/Ki-67(Tex-int 和 Teff)和 TOX/PD-1(Tex-int 和 Tex-term)之間高度協(xié)調(diào)的表達(dá)模式(圖 8E)。 根據(jù)這些標(biāo)準(zhǔn)確定CD8+ Tex-int(元簇 7、10、12、13、18、21 和 22)和 CD8+ Tex-term(元簇17和19)(圖 8C-F)。
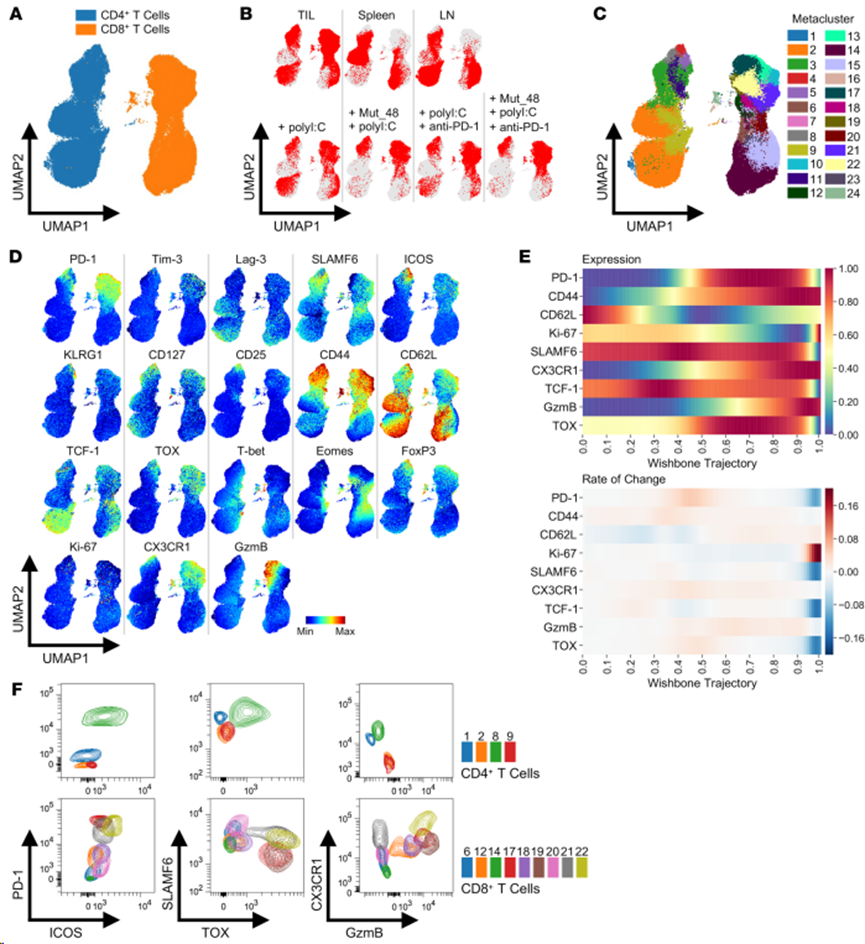
圖8. 腫瘤和外周淋巴器官中CD4+和CD8+ T細(xì)胞亞群的鑒定
接下來(lái),根據(jù)TIL(圖 9A和B)、脾臟(圖 10A和B)和腫瘤引流 Ig LN(圖 10C和D)中各處理間 CD4+ T 細(xì)胞或 CD8+ T 細(xì)胞的頻率對(duì)元集群進(jìn)行解析。在 24 個(gè)元集群中,發(fā)現(xiàn) 4個(gè)CD4+ T 細(xì)胞相關(guān)元集群和 6個(gè)CD8+ T 細(xì)胞相關(guān)元集群相對(duì)于 TIL 中單獨(dú)的polyI:C 處理具有統(tǒng)計(jì)學(xué)意義。在 CD4+ TIL 中,與接種 Mut_48 疫苗相關(guān)的 Tn 和 Teff/mem 細(xì)胞(元簇 1、2 和 9)顯著減少,這與 T 細(xì)胞引誘作用一致。Mut_48 疫苗接種導(dǎo)致 CD4+ Teff/mem 元簇 8 的增加,根據(jù) PD-1 和 ICOS 表達(dá)的增加,這似乎可能是 T 濾泡輔助細(xì)胞(Tfh)的來(lái)源(圖 8F 和圖 9B)。在所有器官和治療中,CD4+Teff/mem(非Treg)沒(méi)有顯示細(xì)胞毒性標(biāo)記(KLRG-1和GzmB),這與它們?cè)谠撃P椭凶鳛檩o助細(xì)胞的作用一致(圖8D和F)。在 CD8+ TIL 中,與 Mut_48 疫苗誘導(dǎo)引物相關(guān)的 Tn(元簇 14)減少。抗 PD-1 治療似乎足以導(dǎo)致 Tprec/prog 亞群(元簇 6 和 20)的擴(kuò)大。單用抗-PD-1(元簇 18)、Mut_48 和抗-PD-1 組合(元簇 12 和 21)或任一處理(元簇 22)都能支持特定的Tex-int 亞群。單獨(dú)接種Mut_48 疫苗后,Tex-term亞群(元簇 17 和 19)被觀察到擴(kuò)大;然而,抗-PD-1 治療阻止這一現(xiàn)象,已知抗-PD-1 會(huì)動(dòng)員 PD-1- Tn 和 PD-1lo Tprec/prog,而犧牲 PD-1hi Tex-term細(xì)胞(圖 8F 和圖 9B)。雖然 Mut_48 和抗 PD-1 聯(lián)合治療支持 TIL 中的 Tprec/prog 分化與單獨(dú)治療相比沒(méi)有差異,但聯(lián)合治療后脾臟和腫瘤引流 Ig LN 中的小群 Tprec/prog(元簇 20)顯著擴(kuò)大(圖 10A-D)。這些數(shù)據(jù)表明,將 PD-1 阻斷與 NeoAg 肽疫苗接種相結(jié)合會(huì)導(dǎo)致非細(xì)胞毒性、輔助性 CD4+ T 細(xì)胞亞群的增長(zhǎng),并更有效地?cái)U(kuò)增外周的 CD8+Tprec/prog 和 TIL 部分中的 Tex-int 群體。
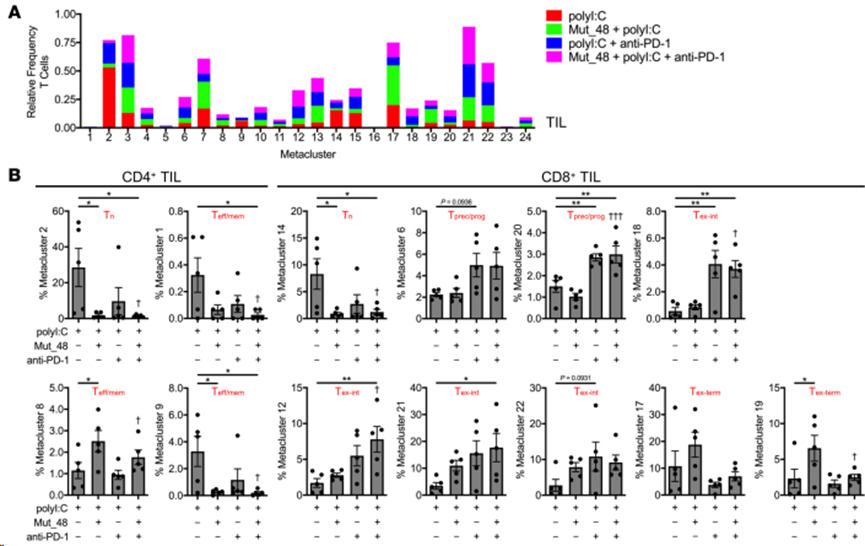
圖9. CltcΔ15疫苗增強(qiáng)抗PD-1誘導(dǎo)的CD8+T細(xì)胞應(yīng)答,并將其重新聚焦于腫瘤中的中間型、效應(yīng)樣亞群
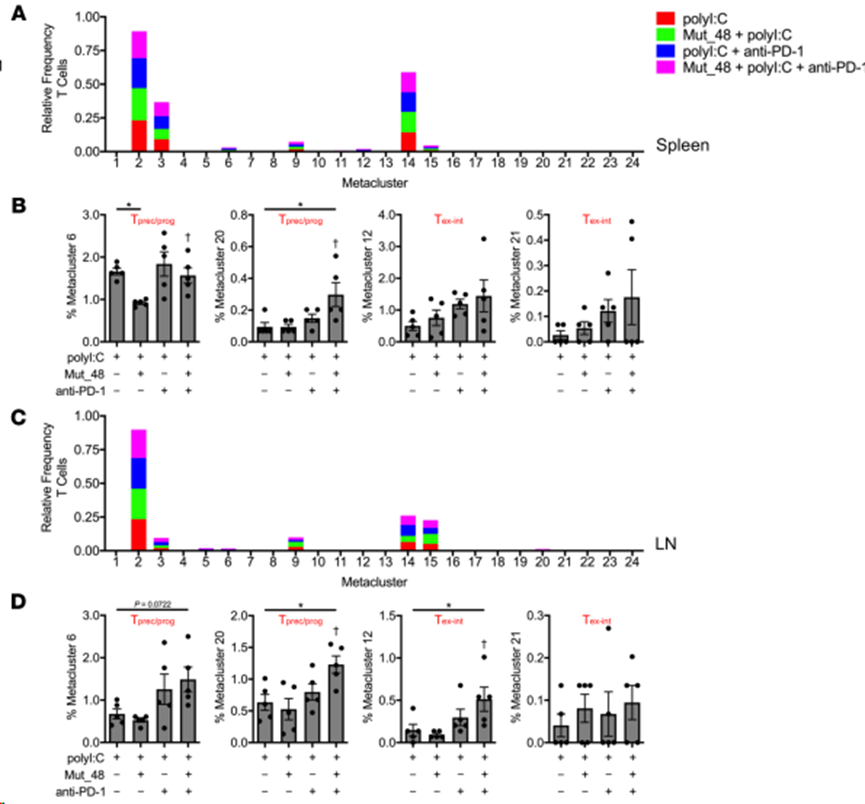
10. 聯(lián)合抗PD-1和CltcΔ15擴(kuò)增外周淋巴器官的前體/祖細(xì)胞和中間耗竭的CD8+T細(xì)胞
5. 新抗原疫苗的療效需要CD4+ T細(xì)胞提供 Th1 型幫助
在接種 NeoAg 肽疫苗前消耗CD4+ T細(xì)胞會(huì)導(dǎo)致部分腫瘤控制,這表明在 CD4+ T細(xì)胞缺失的情況下啟動(dòng)的CD8+ T細(xì)胞并不完全有效,而在治療前消耗 CD8+ T細(xì)胞會(huì)導(dǎo)致腫瘤快速生長(zhǎng),這表明 CD8+ T 細(xì)胞是抗 SCC VII 的必需效應(yīng)因子。在CD4+ T細(xì)胞完全缺失的情況下,激動(dòng)抗CD40交聯(lián)抗體完全恢復(fù)了CD8+ T細(xì)胞介導(dǎo)的腫瘤排斥反應(yīng)(圖11A),這表明即使在CD4+ Tfh缺失的情況下,提供基于Th1的細(xì)胞幫助也是有效治療的關(guān)鍵特征。與此相一致,在疫苗/PD-1阻斷治療聯(lián)合方案中,當(dāng)與 CD8+ T 細(xì)胞最小表位(Mut_48.10)相連時(shí),Cltc CD4+ T 細(xì)胞表位可被 PADRE(X) 替代,并仍能導(dǎo)致完全的腫瘤排斥反應(yīng)(圖 11B)。這些發(fā)現(xiàn)共同表明,功能合理的 NeoAg疫苗聯(lián)合療法可以有效克服不同類型的腫瘤耐藥機(jī)制(無(wú)論是對(duì)化療還是檢查點(diǎn)阻斷單藥)。
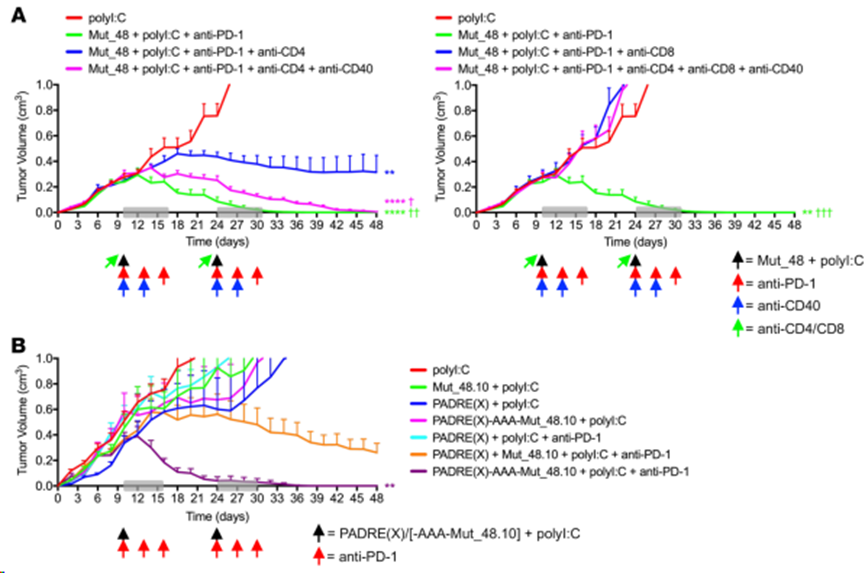
圖11. 栓系CD4+ T細(xì)胞輔助表位通過(guò)CD40依賴機(jī)制優(yōu)化檢查點(diǎn)阻斷和CTL介導(dǎo)的SCCⅶ腫瘤破壞
結(jié)論
綜上所述,本研究證明ICB抗藥性可以通過(guò)與NeoAg疫苗接種相結(jié)合的協(xié)同機(jī)制有效克服,從而維持穩(wěn)定的 CD8+ T 細(xì)胞應(yīng)答,這種應(yīng)答能夠抵御終末衰竭的發(fā)生,并同時(shí)靶向PD-L1+ 和 PD-L1-腫瘤細(xì)胞。可利用本文探索的概念來(lái)開(kāi)發(fā)更有效的個(gè)性化癌癥疫苗,從而擴(kuò)大ICB可治療的腫瘤范圍。
實(shí)驗(yàn)方法
細(xì)胞培養(yǎng),基于腫瘤和肽的免疫和疫苗接種,化療,抗體介導(dǎo)的檢查點(diǎn)阻斷、CD4+/CD8+ T細(xì)胞清除以及對(duì)輔助依賴性的評(píng)估,腫瘤、脾臟和Ig LN單個(gè)核細(xì)胞分離,Q-PCR,WB,ELISPOT,Whole-animal成像,流式細(xì)胞術(shù),光譜數(shù)據(jù)處理與分析,共聚焦顯微鏡,傷口閉合實(shí)驗(yàn),Exome-Seq和RNA-Seq,生物信息學(xué)NeoAg鑒定,生物信息學(xué)分析小鼠與人類RNA-Seq數(shù)據(jù)集的匹配,
參考文獻(xiàn)
Dolina JS, Lee J, Brightman SE, McArdle S, Hall SM, Thota RR, Zavala KS, Lanka M, Ramamoorthy Premlal AL, Greenbaum JA, Cohen EEW, Peters B, Schoenberger SP. Linked CD4+/CD8+ T cell neoantigen vaccination overcomes immune checkpoint blockade resistance and enables tumor regression. J Clin Invest. 2023 Sep 1;133(17):e164258. doi: 10.1172/JCI164258. PMID: 37655661; PMCID: PMC10471175.