






胃癌中METTL16乳酸化通過FDX1 mRNA的m6A修飾促進(jìn)細(xì)胞凋亡

銅是生物體必需的微量元素,參與細(xì)胞生長和代謝。銅穩(wěn)態(tài)失調(diào)與多種腫瘤相關(guān),癌癥患者血清銅水平升高與癌癥分級和化療耐藥相關(guān)。如果銅的濃度超過一定的閾值,就會產(chǎn)生毒性,導(dǎo)致一種新發(fā)現(xiàn)的細(xì)胞死亡形式,即銅中毒。當(dāng)銅直接與三羧酸循環(huán)的脂基化組分結(jié)合時(shí),就會發(fā)生銅死亡,導(dǎo)致隨后含鐵-硫簇的蛋白質(zhì)喪失、蛋白毒性應(yīng)激并最終導(dǎo)致細(xì)胞死亡。鐵氧化還原蛋白1(FDX1)編碼一種還原酶,可將Cu2+還原為毒性更強(qiáng)的Cu1+;硫辛酸二氫硫辛酰胺S-乙酰轉(zhuǎn)移酶(DLAT)是銅中毒所必需的酶。然而,細(xì)胞凋亡在腫瘤中的啟動、傳播和執(zhí)行機(jī)制仍不清楚。m6A是真核細(xì)胞中最豐富的RNA修飾,在腫瘤發(fā)生中起著重要的作用。METTL16在多種腫瘤中以m6A依賴的方式表現(xiàn)出腫瘤發(fā)生和促進(jìn)腫瘤的能力,但控制METTL16活性的調(diào)節(jié)機(jī)制尚不清楚。乳酸源性乳糖化是新近發(fā)現(xiàn)的一種翻譯后修飾(PTM)。乳酸被腫瘤細(xì)胞攝取并轉(zhuǎn)運(yùn)到線粒體進(jìn)行氧化提供能量,同時(shí)衍生組蛋白賴氨酸殘基的乳糖化來刺激基因轉(zhuǎn)錄。組蛋白乳糖化修飾已被證明涉及多個(gè)病理過程,如巨噬細(xì)胞極化和腫瘤發(fā)生。細(xì)胞中非組蛋白的豐度和特異性仍高于組蛋白。非組蛋白是否存在大量的乳糖化修飾,以及這些乳糖化的非組蛋白如何在腫瘤進(jìn)展中發(fā)揮作用和被調(diào)節(jié),這些都亟待探索。該研究發(fā)表在《Nature Communications》,IF:16.6。
技術(shù)路線

結(jié)果
1. 胃癌組織中銅含量較高,且與腫瘤進(jìn)展相關(guān)
銅死亡是由銅濃度過高引起的,但銅死亡在腫瘤中的調(diào)控機(jī)制仍需探索。由于銅主要通過胃和小腸上部吸收,因此胃腸道系統(tǒng)的腫瘤適合研究銅突的發(fā)生和傳播。研究者分析了48對胃癌組織和癌旁組織中的銅濃度,發(fā)現(xiàn)胃癌組織中的銅濃度高于正常胃組織(圖1a)。進(jìn)一步分析發(fā)現(xiàn),ⅲ期胃癌患者的相對銅含量高于ⅰ期和ⅱ期胃癌患者(圖1b),表明銅含量與腫瘤進(jìn)展相關(guān)。此外,銅含量與胃癌患者的總生存期(OS)和無病生存期(DFS)呈負(fù)相關(guān)(圖1c, d)。同時(shí),銅含量與細(xì)胞增殖指標(biāo)Ki-67表達(dá)呈正相關(guān)(圖1e)。同樣,銅含量與患者血清標(biāo)本中的血小板/淋巴細(xì)胞和中性粒細(xì)胞/淋巴細(xì)胞比值呈正相關(guān),兩者都是與炎癥相關(guān)的胃癌侵襲性惡性腫瘤的預(yù)后生物標(biāo)志物(圖1f, g)。這些結(jié)果表明,銅的高含量參與了氣相色譜的發(fā)生和發(fā)展。
此外,研究者還檢測了銅在不同GC類型中的濃度差異,尤其是在惡性腫瘤中。有趣的是,研究者發(fā)現(xiàn)黏液腺癌中的銅濃度高于整個(gè)胃腫瘤(圖1h)。黏液腺癌是胃癌的一種罕見組織學(xué)亞型,與非黏液腺癌相比,黏液腺癌的分期更晚,5年OS和DFS更差。黏液腺癌作為一種難治性胃癌,具有放化療抵抗的特點(diǎn),迫切需要更有效的治療方法。
最近,研究人員發(fā)現(xiàn),細(xì)胞內(nèi)的高銅濃度觸發(fā)了還原酶FDX1,將Cu2+還原為毒性更強(qiáng)的Cu1+,導(dǎo)致銅死亡。研究者發(fā)現(xiàn)FDX1在胃癌中的蛋白水平高于正常胃組織(圖1k)。由于FDX1蛋白水平和銅含量在GC中都較高,因此可能更容易觸發(fā)銅突。這為胃癌尤其是惡性腫瘤——黏液腺癌提供了潛在的治療策略。
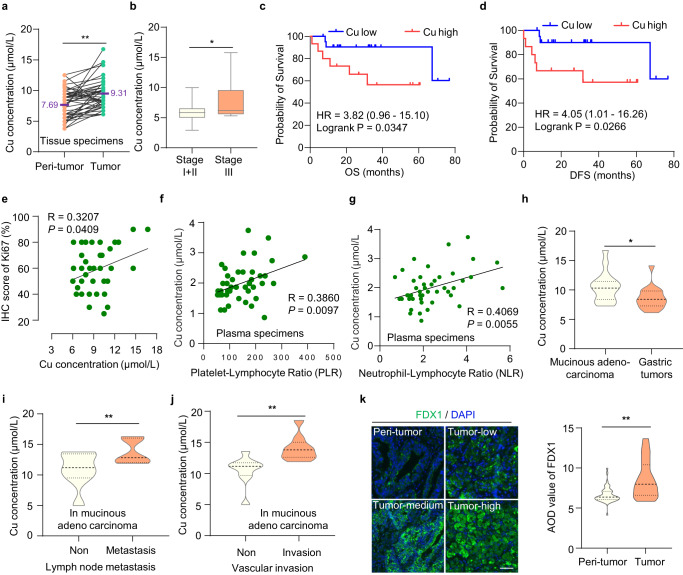
圖1 胃癌組織中銅含量較高,且與腫瘤進(jìn)展相關(guān)
2. METTL16是銅死亡的關(guān)鍵介導(dǎo)因子
有研究表明,與良性胃病患者和健康對照患者相比,胃癌患者的總m6A水平顯著升高,并促進(jìn)胃癌的發(fā)生、生長、侵襲、上皮間質(zhì)轉(zhuǎn)化、轉(zhuǎn)移甚至多藥耐藥的過程。此外,m6A修飾還參與多種細(xì)胞死亡類型,包括凋亡、壞死性凋亡、鐵死亡和焦亡。然而,m6A修飾與銅死亡的關(guān)系尚不清楚。有趣的是,研究者發(fā)現(xiàn)銅含量與GC組織中總m6A水平呈正相關(guān)(圖2a)。此外,銅顯著上調(diào)胃癌細(xì)胞中m6A修飾的總水平(圖2b)。鑒于甲基轉(zhuǎn)移酶樣蛋白(METTL)家族在促進(jìn)m6A修飾中起主導(dǎo)作用,研究者分析了以1:1的比例用來來洛莫/雙硫侖和銅處理METTL敲低的細(xì)胞時(shí)的活力。結(jié)果表明,METTL16敲低導(dǎo)致對來來洛莫/雙硫侖-銅處理的耐藥性(圖2c-e)。此外,在對照組和METTL16敲低的細(xì)胞中,四硫鉬酸鹽處理均抑制了銅綠假單胞菌脫除,表明METTL16參與了銅綠假單胞菌脫除(圖2f)。為了進(jìn)一步驗(yàn)證這些結(jié)果,研究者成功構(gòu)建并驗(yàn)證了METTL16敲除克隆細(xì)胞系,觀察到與METTL16敲除細(xì)胞相似的結(jié)果(圖2g, h)。這些結(jié)果表明,METTL16在銅突病中起著至關(guān)重要的作用。
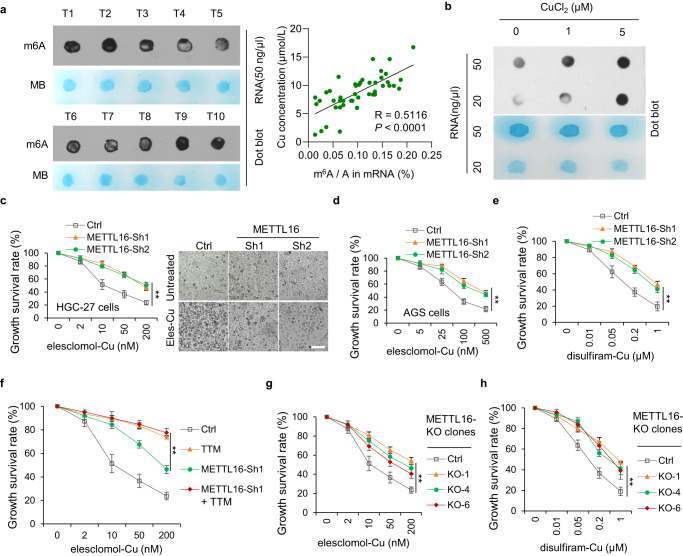
圖2 METTL16是銅死亡的關(guān)鍵介導(dǎo)因子
3. METTL16通過靶向FDX1促進(jìn)銅死亡
METTL16在許多轉(zhuǎn)錄本中負(fù)責(zé)m6A的沉積。因此,研究者在穩(wěn)定的Ctrl-Sh和METTL16-Sh細(xì)胞系中進(jìn)行了甲基化RNA免疫沉淀(MeRIP)測序(圖3a)。使用HOMER軟件預(yù)測每個(gè)樣本中潛在的m6A修飾,m6A修飾主要映射到經(jīng)典的GGAC基序(圖3b)。根據(jù)MeRIP-seq結(jié)果,研究者進(jìn)一步分析了mRNA的總m6A分布模式,發(fā)現(xiàn)m6A峰主要富集在CDS和3'UTR區(qū)域。GO富集分析顯示METTL16修飾基因富集的前15條通路,其中部分與線粒體通路相關(guān)(圖3c)。此外,研究者從METTL16-sh組中鑒定出10個(gè)典型的與m6A甲基化水平低以及mRNA水平下調(diào)相關(guān)的因子(圖3d)。qPCR分析顯示,在METTL16-Sh細(xì)胞中,F(xiàn)DX1、MTF1、PDHB、ATP7A和DLD的mRNA水平降低。與對照細(xì)胞相比,F(xiàn)DX1在METTL16-sh細(xì)胞中表現(xiàn)出最顯著的mRNA水平降低(圖3e)。此外,在METTL16-Sh或-KO細(xì)胞中檢測到FDX1的蛋白水平顯著降低(圖3f, g)。考慮到FDX1是銅離子載體誘導(dǎo)的細(xì)胞死亡的關(guān)鍵調(diào)節(jié)因子,研究者推測METTL16誘導(dǎo)的細(xì)胞凋亡是FDX1依賴的。
接下來,研究者評估了METTL16是否通過影響FDX1的表達(dá)來調(diào)節(jié)細(xì)胞凋亡。鑒于蛋白質(zhì)脂化是銅胞嘧啶的關(guān)鍵因素,研究者使用硫辛酸特異性抗體作為DLAT脂化的指標(biāo),評估了METTL16敲低是否影響蛋白質(zhì)脂化。結(jié)果表明,如免疫印跡測定的那樣,METTL16敲低導(dǎo)致DLAT脂化降低(圖3h)。此外,F(xiàn)DX1過表達(dá)逆轉(zhuǎn)了在來來洛莫/雙硫侖-cu處理后,METTL16敲低和敲除細(xì)胞系中銅突亡的拮抗作用(圖3i, j)。FDX1敲低可抑制METTL16過表達(dá)誘導(dǎo)的銅突(圖3k)。綜上所述,這些數(shù)據(jù)表明METTL16通過靶向FDX1促進(jìn)銅死亡。
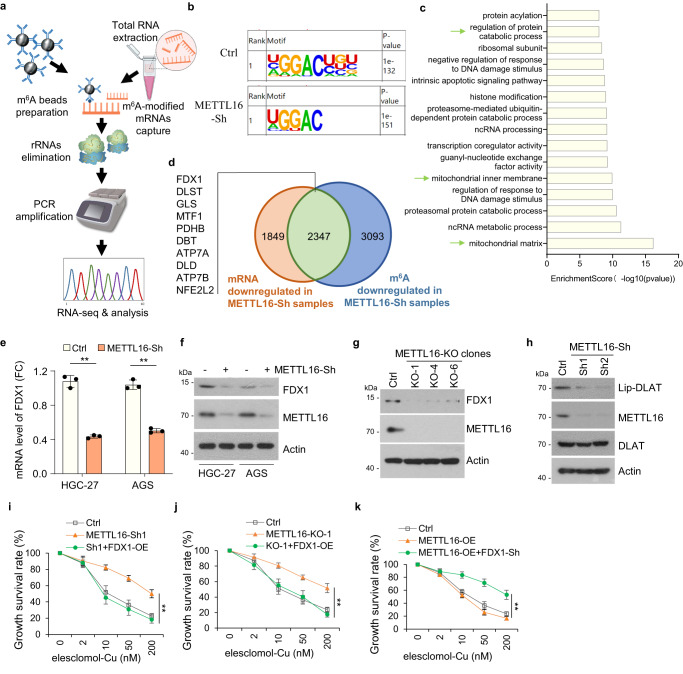
圖3 METTL16通過靶向FDX1促進(jìn)銅突
4. METTL16通過對FDX1 mRNA的m6A修飾促進(jìn)FDX1的積累
為了進(jìn)一步探索METTL16如何調(diào)控m6A修飾進(jìn)而影響FDX1 mRNA水平,研究者進(jìn)行了基因特異性MeRIP-qPCR檢測。研究者發(fā)現(xiàn),在METTL16敲低和敲除細(xì)胞中,F(xiàn)DX1 mRNA上的m6A水平顯著降低,而在轉(zhuǎn)染METTL16過表達(dá)質(zhì)粒的細(xì)胞中,F(xiàn)DX1 mRNA上的m6A水平顯著升高(圖4a, b)。當(dāng)用ActD停止轉(zhuǎn)錄時(shí),METTL16敲低細(xì)胞中FDX1 mRNA的衰減速率比對照細(xì)胞快(圖4c)。這些結(jié)果表明,METTL16介導(dǎo)的甲基化導(dǎo)致FDX1 mRNA的穩(wěn)定性。對FDX1富集的m6A峰的綜合基因組學(xué)觀察(IGV)分析顯示,與對照細(xì)胞相比,METTL16-sh細(xì)胞中的m6A水平降低,表明METTL16可能促進(jìn)FDX1在CDS或3'UTR區(qū)域的m6A修飾(圖4d)。因此,研究者將FDX1的部分CDS和3'UTR區(qū)融合到pGL3熒光素酶報(bào)告基因的下游,構(gòu)建FDX1-WT熒光素酶報(bào)告基因。共轉(zhuǎn)染FDX1熒光素酶報(bào)告基因和不同劑量的METTL16-OE質(zhì)粒,研究者發(fā)現(xiàn)METTL16顯著促進(jìn)FDX1熒光素酶的活性(圖4e)。
為了進(jìn)一步研究,研究者使用基于序列的m6A修飾位點(diǎn)預(yù)測器SRAMP預(yù)測了FDX1 mRNA上可能的m6A修飾位點(diǎn)。綜合圖4e中的預(yù)測結(jié)果和數(shù)據(jù),研究者確定了兩個(gè)潛在的m6A修飾位點(diǎn),它們的置信度非常高:位于CDS區(qū)域的位點(diǎn)602a和位于FDX1轉(zhuǎn)錄本UTR區(qū)域的位點(diǎn)820 A(圖4f, g)。接下來用特異性引物進(jìn)行的MeRIP-qPCR分析證明,F(xiàn)DX1轉(zhuǎn)錄本上的602位點(diǎn)是METTL16介導(dǎo)的甲基化的直接底物(圖4h)。此外,研究者基于FDX1-WT熒光素酶報(bào)告基因,通過將m6A基序中的特異性腺苷(A)替換為胸腺嘧啶(T),設(shè)計(jì)并構(gòu)建了FDX1-602-Mut和FDX1-820-Mut熒光素酶報(bào)告基因(圖4g)。雙熒光素酶實(shí)驗(yàn)結(jié)果表明METTL16不能促進(jìn)602位點(diǎn)突變的FDX1-CDS/UTR報(bào)告載體的熒光素酶活性(圖4i)。GEPIA數(shù)據(jù)庫中METTL16和FDX1的表達(dá)分析表明,METTL16和FDX1在GC中呈正相關(guān)(圖4j)。此外,免疫組織化學(xué)染色顯示,在包含54對胃癌和癌旁正常組織的組織芯片中,METTL16和FDX1的表達(dá)呈正相關(guān)(圖4k)。綜上所述,這些結(jié)果證實(shí)了METTL16介導(dǎo)的FDX1 mRNA的m6A修飾對FDX1 mRNA的穩(wěn)定性至關(guān)重要。
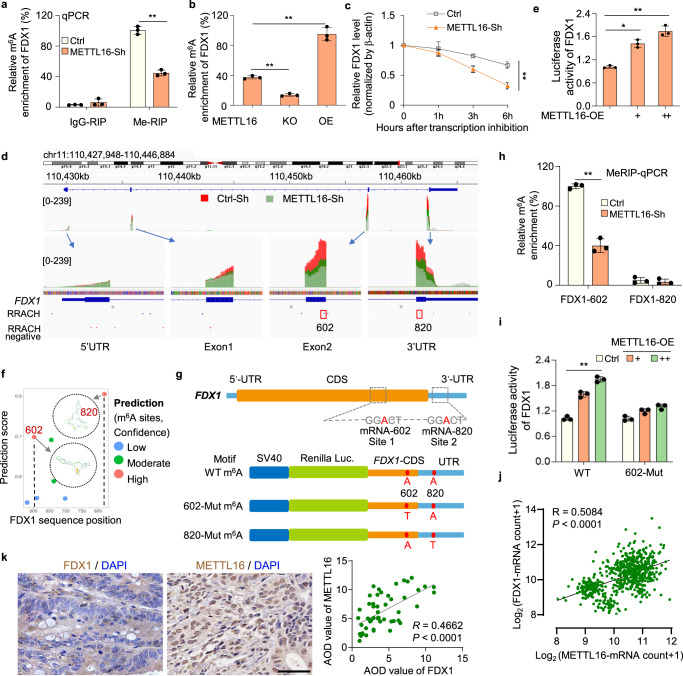
圖4 METTL16通過對FDX1 mRNA的m6A修飾促進(jìn)FDX1的積累
5. 在銅脅迫下,METTL16在K229處發(fā)生乳酸化
接下來,研究者評估了METTL16是否對GC中的銅脅迫有響應(yīng)。在銅處理后,METTL16的mRNA和蛋白水平?jīng)]有變化,但FDX1的蛋白水平顯著增加(圖5a, b)。為了驗(yàn)證在銅脅迫下FDX1表達(dá)的上調(diào)是否依賴于METTL16,研究者檢測了在銅存在或不存在的情況下,METTL16敲低或敲除細(xì)胞中FDX1蛋白的表達(dá)。結(jié)果表明,METTL16缺陷消除了銅誘導(dǎo)的FDX1上調(diào)(圖5c, d)。由于METTL16的mRNA和蛋白水平在銅脅迫下沒有變化,研究者隨后使用質(zhì)譜檢測了METTL16的PTM的變化。分析顯示METTL16蛋白序列上有11個(gè)乳糖化位點(diǎn)和2個(gè)乙酰化位點(diǎn)(圖5e)。在乳酸處理后,6個(gè)乳糖化位點(diǎn)的PTM水平顯著升高,這意味著這6個(gè)位點(diǎn)在腫瘤細(xì)胞中可能受到調(diào)控。然后,研究者證實(shí)了銅脅迫下存在乳糖化或乙酰化反應(yīng)。結(jié)果表明,銅處理顯著增加了METTL16的乳糖化水平而不是乙酰化水平(圖5f)。為了確定METTL16在銅相關(guān)代謝中重要的乳化位點(diǎn),研究者產(chǎn)生了這6個(gè)乳化位點(diǎn)的乳化缺陷突變體(K→R)和K410。研究者發(fā)現(xiàn),只有METTL16-K229R突變體在銅脅迫下顯示出乳糖化降低(圖5g)。隨后,LC-MS/MS分析顯示METTL16-K229與其他乳糖化位點(diǎn)的b-y離子匹配圖見圖5e(圖5h)。為了進(jìn)一步證實(shí)銅脅迫下METTL16-K229的乳糖化,研究者制備了METTL16 lacty-K229抗體,并利用lacty-K229肽段和表達(dá)METTL16-WT、-K229R或-K229E的細(xì)胞驗(yàn)證了其有效性和特異性(圖5i, j)。銅和乳酸處理均誘導(dǎo)K229處METTL16的強(qiáng)乳糖化(圖5k, l),表明K229是銅相關(guān)代謝中必不可少的乳糖化位點(diǎn)。這些結(jié)果表明,在銅脅迫下,METTL16在K229發(fā)生乳酸化。
在穩(wěn)定的METTL16挽救細(xì)胞系中,F(xiàn)DX1蛋白水平進(jìn)一步得到保證,其中內(nèi)源性METTL16被野生型(WT)、乳酸化缺陷型(K229R)或乳酸化模擬型(K229E)取代。結(jié)果顯示,F(xiàn)DX1蛋白水平在METTL16 -敲低的細(xì)胞中降低,在METTL16-WT或-K229E拯救細(xì)胞后恢復(fù),但在METTL16-K229R拯救細(xì)胞后沒有恢復(fù)(圖5m)。銅脅迫誘導(dǎo)METTL16-K229乳糖化,促進(jìn)METTL16甲基轉(zhuǎn)移酶活性。
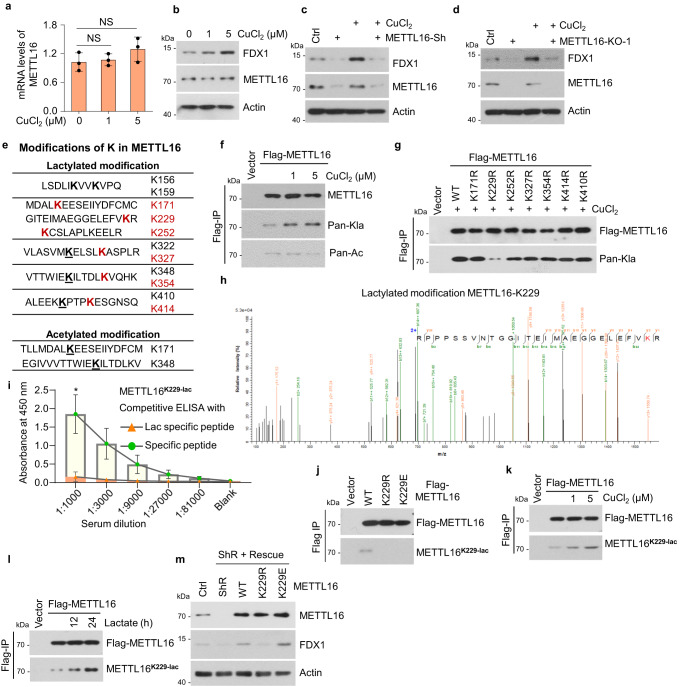
圖5 在銅脅迫下,METTL16在K229處發(fā)生乳酸化
6. SIRT2使METTL16- k229脫酰化并抑制METTL16活性
上述數(shù)據(jù)證實(shí)銅激活METTL16-K229的乳糖化,但具體參與腫瘤細(xì)胞的乳糖化酶和乳糖化轉(zhuǎn)移酶尚不清楚。研究者研究了銅是否介導(dǎo)了對METTL16的乳糖基轉(zhuǎn)移酶或脫乳糖基酶的調(diào)節(jié)。SIRT2被鑒定為METTL16的結(jié)合蛋白,是一種典型的去乙酰化酶,分為主要的脫乙酰酶和乳糖基轉(zhuǎn)移酶組(圖6a)。EIF3b也被鑒定為METTL16的結(jié)合蛋白,這與之前的研究一致。為了研究和驗(yàn)證METTL16和SIRT2之間的相互作用,研究者在HGC-27細(xì)胞中檢測到METTL16-SIRT2的結(jié)合(圖6b)。METTL16-SIRT2的直接關(guān)聯(lián)通過His-pull-down試驗(yàn)得到驗(yàn)證(圖6c)。SIRT2過表達(dá)顯著抑制METTL16-K229的乳糖化(圖6d),而SIRT2敲低或SIRT2抑制劑AGK2處理則促進(jìn)METTL16-K229的乳糖化(圖6e)。此外,銅誘導(dǎo)的METTL16-K229的乳糖化被SIRT2過表達(dá)抑制,而被SIRT2敲低或AGK2處理增加(圖6f, g)。綜上所述,這些結(jié)果表明SIRT2在K229位點(diǎn)與METTL16相互作用并使其脫酰化。
研究者進(jìn)一步探索了SIRT2對METTL16的調(diào)控作用在穩(wěn)定的METTL16挽救細(xì)胞株中的作用。SIRT2過表達(dá)抑制了METTL16- wt細(xì)胞中的FDX1 m6A水平,但未抑制METTL16-K229r或-K229E細(xì)胞中的FDX1 m6A水平,表明SIRT2通過METTL16-K229的乳糖化抑制了METTL16的活性(圖6h, i)。此外,SIRT2過表達(dá)抑制FDX1蛋白水平(圖6j),而SIRT2敲低和AGK2處理促進(jìn)FDX1蛋白水平(圖6k)。免疫熒光染色結(jié)果顯示,在胃癌組織芯片中,SIRT2蛋白水平與FDX1蛋白水平呈負(fù)相關(guān)(圖6l)。綜上所述,這些結(jié)果表明SIRT2通過在K229上去乙酰化來負(fù)調(diào)控METTL16的活性。
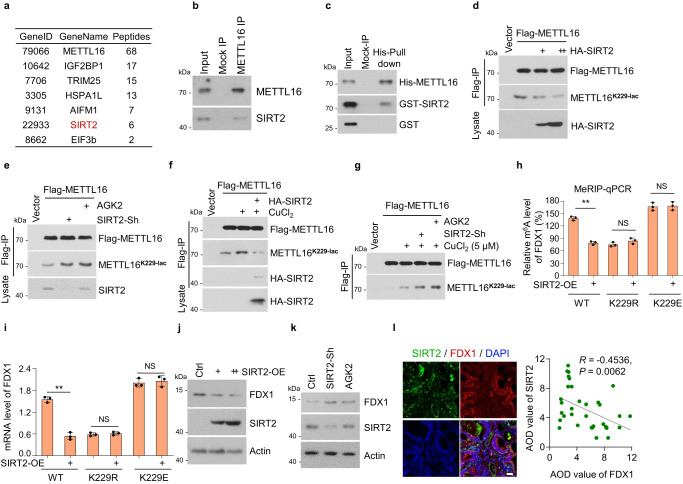
圖6 SIRT2使METTL16-K229脫酰化并抑制METTL16活性
7. SIRT2-METTL16-fdx1細(xì)胞凋亡軸支持一種有前景的治療策略
FDX1是銅死亡的重要介質(zhì),而銅死亡是癌癥的潛在治療方法。研究者發(fā)現(xiàn),在SIRT2抑制或銅誘導(dǎo)METTL16-K229乳糖化后,F(xiàn)DX1蛋白水平升高。接下來,研究者研究了METTL16-K229乳糖化對銅突細(xì)胞的影響。結(jié)果表明,在銅脅迫下,METTL16敲低介導(dǎo)的DLAT脂化降低在METTL16-WT/-K229E細(xì)胞中被恢復(fù),但在METTL16-k229r細(xì)胞中沒有恢復(fù)(圖7a)。此外,在銅存在的情況下,來來洛莫處理可誘導(dǎo)METTL16-WT或-K229E細(xì)胞發(fā)生銅中毒,但對METTL16-K229R細(xì)胞未發(fā)生銅中毒(圖7b)。此外,SIRT2過表達(dá)逆轉(zhuǎn)了在銅脅迫下由METTL16-WT引起的DLAT脂化的增加,而不是由METT16-K229E引起的DLAT脂化的增加,表明SIRT2通過去乙酰化METTL16-K229來抑制銅死亡(圖7c)。AGK2處理同樣促進(jìn)了銅脅迫下METTL16-WT細(xì)胞的DLAT脂酰化,但對METTL16-K229R細(xì)胞沒有作用(圖7d)。此外,AGK2處理促進(jìn)了在有來昔洛莫和銅存在的情況下的銅突(圖7e)。
研究者進(jìn)一步評估了METTL16乳糖化是否可以增強(qiáng)接受來來洛莫治療的小鼠異種移植瘤的治療反應(yīng)。將METTL16-WT、-K229R、-K229E細(xì)胞分別于裸鼠左右后肢上方皮下注射。在METTL16-WT和-K229E組中,接受來來洛莫治療的小鼠的腫瘤生長明顯受到抑制,而在METTL16-K229R組中,腫瘤的體積和重量減少(圖7f-h)。
腫瘤切片中的FDX1染色顯示,METTL16-WT和-K229E組的腫瘤切片中FDX1染色高于METTL16-K229R組(圖7i)。這些結(jié)果表明,METTL16在K229的泌乳狀態(tài)在決定銅突的發(fā)生中起著至關(guān)重要的作用。此外,在異種移植瘤中使用來來洛莫和AGK2聯(lián)合治療。對腫瘤切片的分析表明,AGK2在體內(nèi)可致敏來來洛莫治療,如觀察到的腫瘤體積和重量的減少(圖7j-l)。這些結(jié)果表明,來來洛莫聯(lián)合AGK2治療胃癌,尤其是惡性腫瘤黏液腺癌是一種有前景的治療方法。
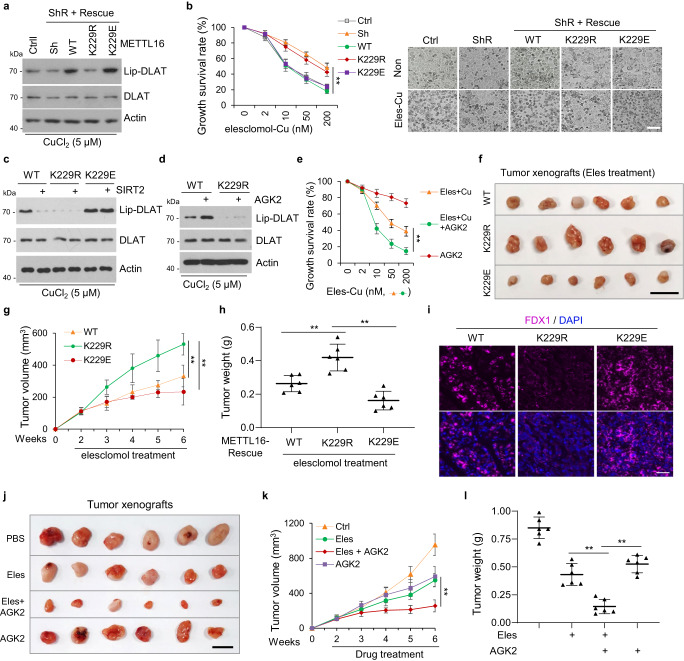
圖7 SIRT2 - METTL16 - fdx1細(xì)胞凋亡軸支持一種有前景的治療策略
結(jié)論
這些結(jié)果表明,METTL16是銅死亡的驅(qū)動力。METTL16在K229上的乳化通過對FDX1 mRNA的m6A修飾上調(diào)FDX1蛋白的表達(dá),這是銅脅迫下觸發(fā)銅中毒的途徑之一。此外,研究結(jié)果表明METTL16在胃癌中是協(xié)調(diào)葡萄糖和銅代謝的中心樞紐。此外,結(jié)果表明METTL16的乳糖化為評價(jià)銅離子載體藥物的有效性提供了一個(gè)潛在的靶點(diǎn)。鑒于胃腫瘤比正常組織具有更高的銅和乳酸濃度,利用銅離子載體結(jié)合銅離子和SIRT2特異性抑制劑可能是一種可行的胃癌治療策略。
實(shí)驗(yàn)方法
細(xì)胞培養(yǎng),小鼠異種移植腫瘤實(shí)驗(yàn),m6斑點(diǎn)印跡測定,qRT-PCR,熒光素酶報(bào)告基因檢測,蛋白質(zhì)印跡,His-pull-down測定,免疫沉淀,免疫熒光染色,MeRIP-qPCR,RNA穩(wěn)定性檢測,體外乳酸化試驗(yàn),生物信息學(xué)分析
參考文獻(xiàn)
Sun L, Zhang Y, Yang B, Sun S, Zhang P, Luo Z, et al. Lactylation of METTL16 promotes cuproptosis via m6A-modification on FDX1 mRNA in gastric cancer. Nat Commun. 2023 Oct 20;14(1):6523. doi: 10.1038/s41467-023-42025-8.