






調節核糖體生物發生——TERT加速BRAF突變誘導甲狀腺癌去分化與進展
欄目:最新研究動態
發布時間:2024-01-23
TERT再激活經常發生在人類惡性腫瘤中,尤其是晚期癌癥。然而,TERT再激活在癌癥進展中的體內功能及其潛在機制尚不完全清楚......
TERT再激活經常發生在人類惡性腫瘤中,尤其是晚期癌癥。然而,TERT再激活在癌癥進展中的體內功能及其潛在機制尚不完全清楚。在本研究中,作者在小鼠甲狀腺上皮中特異性表達了TERT和/或活性BRAF (BRAF V600E)。雖然BRAF V600E單獨誘導乳頭狀甲狀腺癌(PTC),BRAF V600E和TERT共同表達導致低分化甲狀腺癌(PDTC)。空間轉錄組分析顯示,共表達BRAF V600E和TERT的小鼠腫瘤具有高度異質性,細胞去分化與核糖體生物發生呈正相關。從機制上講,TERT通過與參與核糖體生物發生的多種蛋白質相互作用,促進核糖體RNA (rRNA)的表達和蛋白質合成。此外,作者發現CX-5461是一種rRNA轉錄抑制劑,能有效阻斷甲狀腺癌的增殖并誘導其再分化。因此,TERT通過誘導癌細胞去分化來促進甲狀腺癌進展,而核糖體抑制是治療TERT再激活癌癥的潛在策略。本文于2023年8月發表在《Science Advances》IF: 13.6期刊上。
技術路線

主要實驗結果
1、TERT再激活在多種癌癥中經常發生,預示著較差的預后
由于TERT啟動子沒有被全外顯子測序所覆蓋,TERT再激活在癌癥基因組學研究中被低估。最近,隨著TERT啟動子C228T和C250T突變的發現,TERT啟動子狀態在幾項大規模泛癌癥測序研究中得到檢驗。MSK-IMPACT、MSK-MET和China Origimed2020三個包括最豐富的癌癥類型和樣本的大型隊列,分析了TERT啟動子區域。考慮到TERT再激活主要通過啟動子突變或基因擴增發生,作者掃描了這三個隊列中的兩種事件,以及癌癥中的TERT擴增基因組圖譜計劃(TCGA)隊列。作者發現TERT啟動子突變(C228T或C250T)在甲狀腺癌、黑色素瘤、膀胱癌、肝癌、膠質瘤和頭頸癌中經常發生,而TERT擴增在肺癌、卵巢癌和膀胱癌中常見(圖1A)。在不同的癌癥類型中,黑色素瘤和甲狀腺癌在不同的隊列中表現出高度的一致性。此外,在紀念斯隆·凱特琳癌癥中心(MSKCC)數據集顯示,TERT啟動子突變預測甲狀腺癌、膠質瘤和膀胱癌的預后較差(圖1B)。
作者還研究了503個細胞系,這些細胞系通過全基因組測序和/或靶向測序在癌細胞系百科全書(CCLE)數據集中定義了TERT啟動子狀態(24)。啟動子突變是甲狀腺癌和黑色素瘤中TERT激活的主要改變。在這兩種癌癥類型中,來自轉移灶的細胞系顯示出更高頻率的TERT啟動子突變(圖1C),表明TERT激活在癌癥進展和轉移中的潛在功能。
TERT啟動子突變(C228或C250)的頻率與甲狀腺癌的病理分期和分化狀態有關。分化型甲狀腺癌與良好的生存率相關,TERT啟動子突變的發生頻率為4%至12%。同時,約50%預后不良的低分化甲狀腺癌(PDTC)和間變性甲狀腺癌(ATC)存在TERT啟動子突變(圖1D)。值得注意的是,大多數TERT啟動子突變的腫瘤也表現出BRAF或NRAS突變(圖1E)。TCGA計劃建立了甲狀腺分化評分(TDS)系統,通過16個基因(TG, TPO, PAX8, DIO1, DIO2, DUOX1, DUOX2, FOXE1, GLIS3, NKX2-1,SLC26A4, SLC5A5, SLC5A8, THRA, THRB, TSHR)的mRNA表達水平評估甲狀腺癌分化水平。表達分析顯示,在CCLE甲狀腺癌細胞系中,TERT表達水平與TDS評分呈負相關(R = - 0.73, Spearman P = 0.021)(圖1F)。總之,這些結果表明,TERT再激活,特別是啟動子突變,在各種癌癥中經常發生,并與腫瘤進展和不良預后相關。

圖1 TERT再激活預示著更糟糕的預后
2、TERT加速BRAF v600e誘導的甲狀腺癌進展
如上所述,TERT啟動子突變通常與BRAF或NRAS激活突變同時發生(圖1E)。作者推測,TERT過表達可能會促進已有致癌信號的癌癥進展,而不是啟動癌變。先前的研究表明,TERT在體外誘導增殖、侵襲和許多其他腫瘤樣行為。
為了闡明體內TERT再激活與腫瘤進展之間的潛在機制,作者通過將Loxp -stop- loxp- Tert - 3xFlag - IRES - EGFP序列插入Rosa26等位基因(RsmTERTLSL/+小鼠;圖1G)建立了條件TERT轉基因小鼠模型。然后將小鼠與TPOCreERT2小鼠雜交,產生甲狀腺特異性Tert轉基因小鼠模型(TC小鼠),該模型模擬腫瘤中TERT的高表達。然后,作者采用先前使用的甲狀腺癌小鼠模型,通過在甲狀腺濾泡細胞(BC小鼠)中表達BRAF V600E誘導,與Rs-mTERTLSL/+小鼠雜交,生成BTC小鼠(圖1G)。
隨訪超聲顯示,在誘導后的前7 ~ 10個月,BC和BTC甲狀腺腫瘤的生長模式無顯著差異(圖1,H ~ J)。然而,在10至14個月后,BTC小鼠的甲狀腺腫瘤生長速度遠快于BC小鼠(圖1,H至J)。BTC和BC小鼠的腫瘤進展也通過18F-FDG正電子發射斷層掃描-計算機斷層掃描(PET-CT)進行了分析。12個月BTC甲狀腺腫瘤的最大標準化攝取值(SUVmax)遠高于12個月BTC和8個月BTC小鼠,而12個月的BC和8個月的BTC小鼠FDG SUVmax相似(圖1K)。與上述結果一致,BTC組預后較差,中位生存期為406天,而BC組為486.5天(圖1L)。總之,這些結果表明,當BRAF和TERT同時被激活時,腫瘤進展更快。
3、TERT促進BRAF v600e誘導的甲狀腺癌的去分化
與BC小鼠相比,BTC小鼠腫瘤生長加快,預后更差(圖1,J和L)。作者隨后表征了不同階段腫瘤的形態和分子變化。超聲檢查、解剖后的大體外觀、hematoxylin和伊紅(H&E)病理顯示,大多數BTC腫瘤進展為10 ~ 14個月出現較大的PDTC(15 / 17, 88.2%)。與此形成鮮明對比的是,盡管一些老年BC小鼠(16 - 18個月)的腫瘤進展為PDTC(5 / 14, 35.7%),但大多數BC甲狀腺腫瘤仍然是乳頭狀甲狀腺癌(PTCs)(圖2A)。免疫組化(IHC)結果顯示,BC小鼠腫瘤中甲狀腺癌標志物高表達,包括甲狀腺球蛋白(TG)、甲狀腺轉錄因子-1(TTF-1)、配對盒8(PAX8)、鈉碘同向轉運體(NIS;Slc5a5)和CK19。這些標志物已廣泛用于臨床甲狀腺癌的鑒定。然而,TG和TTF-1在BC腫瘤中的表達低于鄰近的正常濾泡細胞(圖2A)。另一方面,BTC小鼠的腫瘤表達低水平的TG、TTF-1、NIS和PAX8,高水平的Ki67,表明過度增殖和去分化(圖2A)。從異位Flag-TERT的表達可以看出,BTC小鼠的腫瘤起源于甲狀腺濾泡細胞(圖2A)。
晚期甲狀腺癌經常轉移到遠處器官,包括肺。作者觀察到BTC肺內廣泛的轉移灶(圖2B)。轉移灶細胞SP-C(AT2肺細胞的標志物)和TG呈陰性,TTF-1呈陽性,PAX8和NIS呈弱陽性,表明這些轉移細胞起源于甲狀腺(圖2C)。
作者分析BC和BTC腫瘤的轉錄組,以確定細胞去分化和腫瘤轉移的潛在機制。收集BC和BTC小鼠(n = 4和5)的腫瘤,進行RNA測序(RNA-seq)。主成分分析表明BTC和BC甲狀腺腫瘤的基因表達譜明顯不同(圖2D)。整體評估顯示BTC腫瘤的TDS評分要低得多(圖2E)。在BTC腫瘤中,甲狀腺特異性基因如Tg、Pax8、Tshr和Tpo下調,而多種腫瘤基因如Lgr5、Clu、Mmp9、Pappa2、Mki67和Arg1上調(圖2F)。作者能夠從BTC腫瘤(312BTC)中分離和永生化原代腫瘤細胞。這些結果與免疫組化分析一致,表明BTC小鼠腫瘤具有高增殖和去分化表型。
為確定與BTC小鼠從PTC到PDTC進展相關的關鍵癌癥途徑,對標記基因集進行基因集富集分析(GSEA)和基因集變異分析(GSVA)。幾個癌癥標志,如E2F靶點、G2-M檢查點、上皮-間質轉化、炎癥反應、程序性細胞死亡1配體1(PD-L1)表達和程序性細胞死亡蛋白1(PD-1)檢查點通路,在BTC腫瘤中被激活,而氧化磷酸化和脂肪酸代謝則被下調(圖2、G、H)。此外,KEGG富集分析差異表達基因表明,細胞因子-細胞因子受體相互作用、磷脂酰肌醇3-激酶(PI3K)-AKT信號通路、白細胞介素-17(IL-17)信號通路、細胞外基質(ECM)-受體相互作用和腫瘤壞死因子(TNF)信號通路在BTC腫瘤中富集(圖2I)。
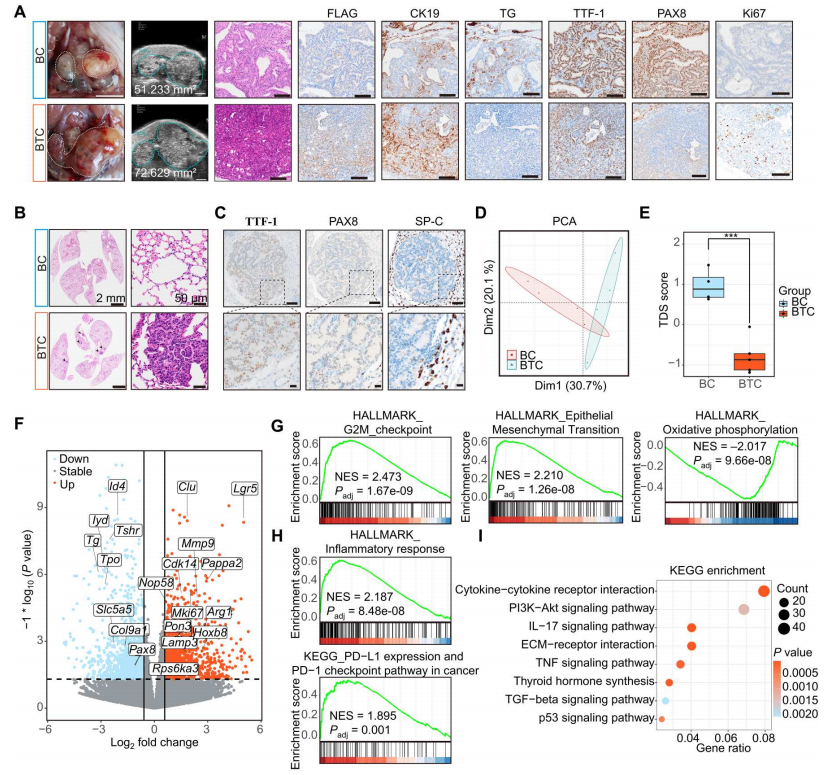
圖2 TERT促進BRAF突變誘導的甲狀腺癌的去分化
4、BTC腫瘤的空間轉錄組分析揭示了rRNA代謝相關的去分化
為了進一步探討TERT在BRAF V600e誘導的甲狀腺癌去分化中的作用,作者對來自BTC小鼠的具有高度異質性的腫瘤進行了Visium 10X空間轉錄組(ST)分析。腫瘤具有正常濾泡細胞、PTC和PDTC的特征區域,可能代表了甲狀腺癌的整個發展軌跡(圖3A)。
為探索甲狀腺細胞的轉錄異質性,作者重新聚集了637個甲狀腺上皮斑點(圖3B和C)。簇0(c0)中的細胞表達高水平的Tg和Tpo,標志著分化良好的濾泡細胞(圖3D)。一些TDS特征基因如Slc5a5、Thrb、Duox1、Duox2和Tshr在ST分析中未檢測到;因此,作者定義一個以Tg、Tpo、Dio1、Fhl1和Sorbs2為特征基因的mST_TDS(小鼠ST TDS)。mST_TDS評分顯示,c0是分化較好的聚類,其次是c1和分化較差的c2/c3/c4/c5(圖3E)。
為了追蹤甲狀腺簇的進化動態,作者使用Monocle3進行了軌跡分析,并將分化良好的濾泡細胞c0作為軌跡起始根(圖3F)。在UMAP投影顯示,甲狀腺細胞表現出“c0-c1/c2/c3/c4-c5”的進化模式,其中c0和c5分別收集在軌跡路線的起點和終點,其他四個簇分布在兩者之間(圖3G)。因此,作者可以假設甲狀腺細胞從c0(表示為state1)進化,通過中間簇c2/c3/c4(state2),最終去分化為c5(state3)。為進一步確定分子途徑的改變,使用AUCell計算三種狀態的Reactome富集分數(圖3H)。與c0正常甲狀腺表現一致,state1富集于甲狀腺素生物合成途徑。State2呈現介于state1和state3之間的中間狀態,主要富集ECM和免疫相關通路。有趣的是,state3在端粒維持中比stat1/2更活躍,并且在rRNA加工和翻譯途徑中富集,這反映了核糖體的生物發生和活性(圖3H和I)。此外,細胞對缺氧和饑餓的反應等物理應激也在state3細胞中豐富(圖3H)。這一觀察結果表明,核糖體生物發生在c05/state3細胞中發揮重要作用。
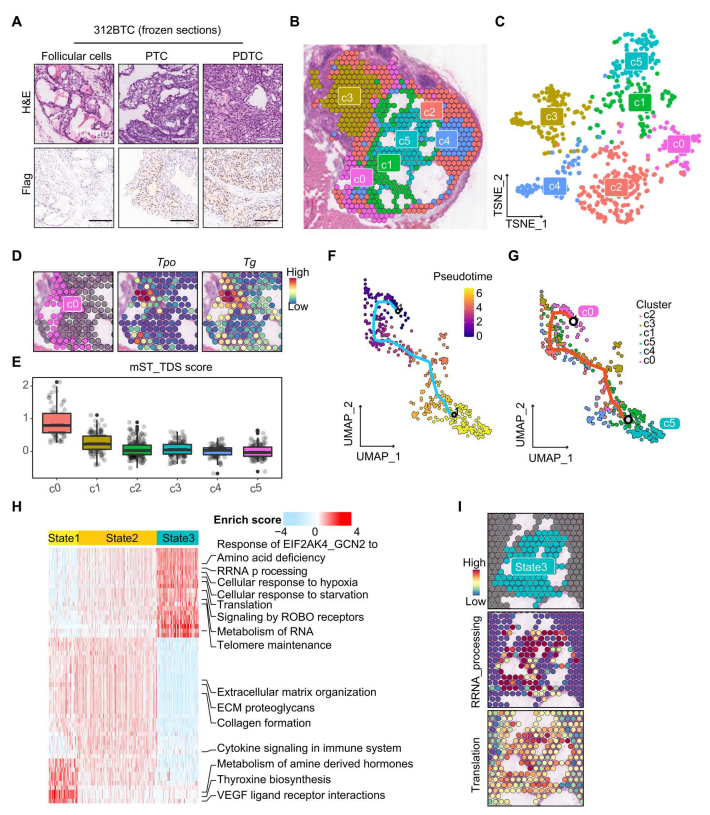
圖3 空間轉錄組分析揭示了BTC小鼠甲狀腺的異質性
5、TERT誘導rRNA合成和核糖體活性
細胞外信號調節激酶(ERK)激活被認為是rRNA轉錄的上游信號,可能通過上游結合轉錄因子(UBF)的磷酸化。此外,TERT已被證明可以誘導rRNA和tRNA轉錄。作者評估NTHYCTL、NTHY-B(BRAF V600E)和NTHY-BT(BRAF V600E)細胞中pre-45SrRNA水平。作者發現BRAF V600E上調pre-45SrRNA轉錄,并進一步被TERT誘導。因此,TERT和BRAF V600E可能共同促進核糖體的生物發生。
在癌細胞中,核糖體經常失控,導致不受控制的增殖和轉移。探索TERT再激活是否與癌癥中核糖體活性相關,作者分析CCLE數據集中與TERT共表達的基因。與TERT表達呈正相關的基因(Spearman R > 0.25, P < 0.01)參與細胞周期進程和rRNA代謝(圖4A)。核糖體由rRNA和核糖體蛋白組成,核糖體的生物發生需要所有RNA聚合酶(Pol I/II/III),以及小核RNA(snoRNA)和參與rRNA過程和成熟的多種其他酶的參與。作者發現在CCLE數據庫中TERT與rRNA轉錄調控因子UBTF、POLR3E和FBL以及rRNA外切酶EXOSC2呈正相關(圖4B)。然后作者分析了最近的甲狀腺癌單細胞RNA-seq(scRNA-seq)數據集。由于在scRNA-seq中沒有有效檢測到TERT,因此作者使用Reactome_端粒維持富集評分作為TERT活性的替代品。在甲狀腺細胞(n = 42,708個細胞)中,Reactome_端粒維持富集評分與rRNA加工(R = 0.53, P < 2.2 × 10?16)和翻譯(R = 0.56, P < 2.2 × 10?16)呈正相關(圖4C)。這些結果表明TERT與核糖體代謝密切相關。
作者進一步檢查TERT對不同癌細胞中rRNA的影響。在K1(甲狀腺癌)、OCM1(黑色素瘤)和BCPAP(甲狀腺癌)細胞(均有TERT啟動子C228T或C250T突變)中TERT敲低后,觀察到rRNA水平急劇下降(圖4D至F)。rRNA的下調不僅發生在Pol I轉錄的18S、5.8S和28S中,也發生在Pol Ⅲ轉錄的5S rRNA中(圖4E)。相反,異位TERT在甲狀腺細胞系MDA-T41和結腸癌細胞系RKO和HT-29[均具有野生型(WT) TERT啟動子]中顯著誘導rRNA表達(圖4F)。由于80%以上的細胞RNA是rRNA,因此采用乙基尿苷(EU)法標記新生RNA,尤其是rRNA。在K1和OCM1細胞中,TERT敲低顯著抑制細胞核新生rRNA合成,而纖維蛋白(FBL;Cajal體成分)沒有改變(圖4G到J)。總之,這些結果表明TERT是最佳rRNA合成所必需的。
由于TERT是端粒酶的主要亞基之一,作者想知道端粒酶活性是否參與rRNA的合成。作者發現,在BCPAP細胞中,TERC(端粒酶的另一個主要成分)的敲低導致rRNA表達減少(圖4K),表明TERT對rRNA合成的調控至少部分依賴于端粒酶活性。
核糖體的主要功能是翻譯,以mRNA為模板合成蛋白質。由于TERT可以增強rRNA轉錄,作者想知道TERT是否可以調節翻譯過程。在作者的ST數據和CCLE數據集中,高TERT表達與翻譯相關(圖3H和4L)。為了驗證TERT在翻譯中的作用,作者采用了兩種方法來檢驗翻譯效率。有趣的是,表面翻譯感知(SUnSET)實驗表明,TERT敲低導致BRAF和TERT啟動子突變的K1和SW1736細胞中新生蛋白的產生減少(圖4M)。在雙順反子報告實驗中,TERT的敲低抑制了5’帽和內部核糖體進入位點(IRES)依賴的翻譯效率(圖4N)。因此,在TERT再激活的癌細胞中,蛋白質合成似乎依賴于TERT。

圖4 TERT誘導rRNA合成和核糖體活性
6、TERT與核糖體支架相互作用并增強MTORC1活動
功能性端粒酶復合體除TERT和TERC外,還包括DKC1和核糖體生物發生因子1(PES1)等多種組分。DKC1和PES1與核糖體功能有關。因此,作者探究TERT是否通過與rRNA代謝相關蛋白相互作用參與核糖體途徑。作者采用免疫沉淀和質譜法來鑒定TERT的潛在結合伙伴。已經確定參與RNA分解代謝過程、核糖體生物發生、翻譯控制、rRNA加工和端粒維持的幾種蛋白質(圖5A)。此外,TERT與參與酵母和人pre-rRNA加工的Bystin Like (BYSL)之間的相互作用在人胚胎腎(HEK) 293T和甲狀腺癌細胞K1中都被鑒定出來(圖5B)。此外,作者在K1、HEK293T和小鼠原發性甲狀腺癌細胞系312BTC中驗證TERT與BYSL以及EIF2亞基之間存在蛋白相互作用(圖5C)。總的來說,這些結果表明TERT可能通過與已知的核糖體調節劑相互作用來調節核糖體的生物發生。
PI3K-AKT和MTORC1信號通路在調節核糖體的蛋白質合成中起主要作用。在作者的整體和空間轉錄組分析中,PI3K-AKT和MTORC1信號通路在BTC腫瘤中富集(圖5D)。與此一致的是,參與MTORC1通路的多個基因,如Rpn1、Xbp1、Eif2s2,在BTC組中上調(圖5E)。此外,甲狀腺癌的scRNA-seq數據顯示,端粒維持與甲狀腺細胞中MTORC1介導的信號傳導存在顯著相關性(R = 0.43, P < 2.2× 10?16)(圖5F)。此外,作為MTORC1激酶活性的讀數,p-S6水平在BTC小鼠腫瘤中顯著增強(圖5G)。為了進一步研究TERT在甲狀腺癌MTORC1信號傳導中的作用,作者在MDA-T41細胞中表達WT-TERT或DN-TERT(無端粒酶活性的突變體),并分析MTORC1已知下游效應物的磷酸化(圖5H和I)。與體內實驗結果一致,WT-TERT的表達有效地誘導了S6和S6K的磷酸化,特別是在血清饑餓條件下(圖5I和J)。因此,MTORC1也可能參與TERT對核糖體的激活。

圖5 TERT與核糖體支架相互作用,增強MTORC1活性
7、靶向抑制rRNA轉錄可阻斷腫瘤進展
TERT下游的靶向機制,如核糖體生物發生和MTORC1信號傳導,可能作為與TERT抑制劑相比副作用更小的替代抗腫瘤策略。作者分析癌癥藥物敏感性基因組學(GDSC)數據集,發現TERT表達水平較高的細胞對CX-5461更敏感,CX-5461是一種Pol Ⅰ介導的rRNA轉錄抑制劑,在正常體細胞中耐受。CX-5461呈現出與之前的端粒酶靶向藥物如BIBR 1532和端粒酶抑制劑IX類似的模式(圖6A)。采用BRAF V600E和TERT啟動子共突變體BCPAP和OCM1細胞以及小鼠312BTC細胞檢測CX-5461的作用。CX-5461以劑量依賴的方式抑制癌細胞增殖(圖6B至D)。隨后建立BCPAP, OCM1和小鼠611BTPC(Trp53失活的312BTC細胞)皮下腫瘤模型,并口服CX-5461在體內檢測CX-5461的抗腫瘤作用。作者發現CX-5461在所有三種模型中都有效地抑制了腫瘤生長,并且在所測試的小鼠中沒有或輕微的體重減輕(圖6E至J)。此外,在CX-5461治療后,腫瘤中Ki67的表達也降低了(圖6K至C)。綜上所述,這些結果表明CX-5461能夠有效抑制BRAF突變和TERT再激活細胞的增殖。
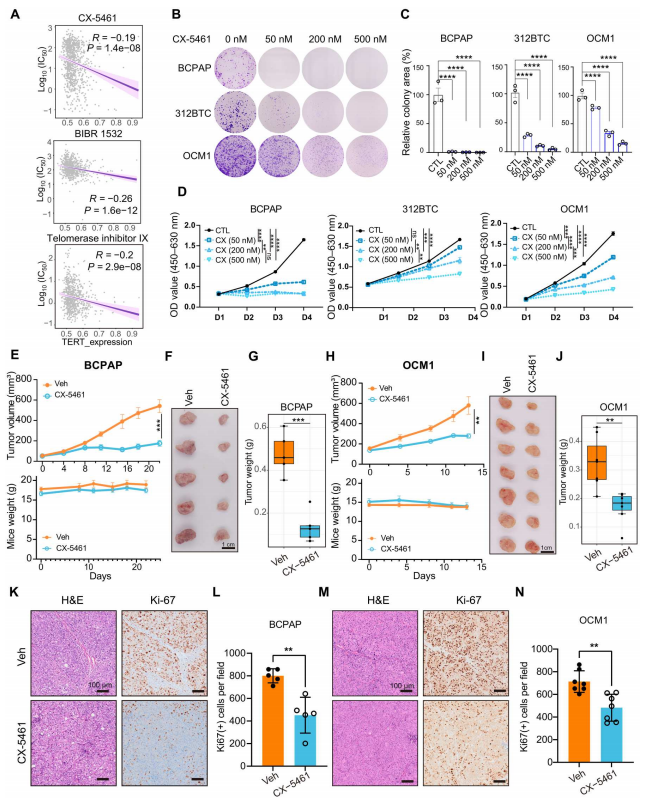
圖6 CX-5461抑制多發性腫瘤進展
8、CX-5461誘導甲狀腺癌再分化
放射性碘(RAI)治療被認為是轉移性甲狀腺癌的一線治療方法。然而,幾乎所有的PDTC和ATC,以及一些分化良好的甲狀腺癌,由于功能性甲狀腺濾泡標志物尤其是碘轉運蛋白NIS的表達減少,不能有效地吸收碘。這些患者被歸類為RAI難治性,而難治RAI是甲狀腺癌相關死亡的主要原因之一。鑒于TERT在腫瘤進展中的主要作用是誘導腫瘤去分化,阻斷TERT或其下游效應物可能導致腫瘤分化。
因此,作者評估CX-5461對甲狀腺癌細胞分化標志物的影響。在CX-5461處理24或48小時后,K1、BCPAP和SW1736細胞中PAX8、DIO1、THRA、THRB、FOXE1和TSHR的表達水平明顯上調(圖7A至C)。CX-5461治療后,K1和SW1736細胞中的NIS和PAX8蛋白表達水平也升高(圖7D至F)。隨后,作者測試CX-5461治療是否可以增加甲狀腺腫瘤的碘攝取。采用611BTPC同基因腫瘤模型檢測CX-5461給藥前后腫瘤碘攝取的差異。結果顯示,CX-5461治療誘導了皮下611BTPC腫瘤的碘攝取,而原生甲狀腺的碘攝取保持不變(圖7G和H)。綜上所述,rRNA轉錄阻斷是誘導甲狀腺癌再分化的有效途徑。

圖7 CX-5461誘導甲狀腺癌再分化
結論
作者在本研究中證明了TERT的條件轉基因表達促進了小鼠甲狀腺BRAF V600E突變驅動的去分化和腫瘤轉移。TERT在小鼠和人BRAF V600E腫瘤細胞中的表達誘導rRNA轉錄和核糖體生物發生。rRNA轉錄抑制劑CX-5461誘導腫瘤分化,減緩腫瘤進展(圖8)。

圖8 研究示意圖
綜合結合動物模型、ST測序、數據庫分析和實驗揭示了核糖體途徑中的TERT。
實驗方法
細胞培養;建立小鼠甲狀腺原代細胞系;質粒和慢病毒包;質粒和siRNA轉染;動物超聲成像和PET-CT;活體[125I]攝取實驗;免疫組織化學染色;RNA提取,cDNA合成和RT-qPCR;Bulk RNA-seq;ST樣品制備,文庫構建和測序;ST數據處理和可視化;軌跡分析;甲狀腺癌單細胞數據集分析;單細胞或ST富集分析;EU assay;免疫印跡;免疫沉淀;液相色譜-質譜;端粒重復擴增實驗;表面翻譯感知;雙鏈雙熒光素酶報告基因;集落形成;細胞計數試劑盒;甲狀腺癌患者OncoVee MiniPDX
參考文獻
Yu P, Qu N, Zhu R, Hu J, Han P, Wu J, Tan L, Gan H, He C, Fang C, Lei Y, Li J, He C, Lan F, Shi X, Wei W, Wang Y, Ji Q, Yu FX, Wang YL. TERT accelerates BRAF mutant-induced thyroid cancer dedifferentiation and progression by regulating ribosome biogenesis. Sci Adv. 2023 Sep; 9(35): eadg7125. doi: 10.1126/sciadv.adg7125.