






m5C甲基化lncRNA NR_033928促進谷氨酰胺代謝重編程從而促進胃癌增殖

5-甲基胞嘧啶(m5C)異常甲基化已被證實與胃癌發生、發展及預后密切相關。失調的長鏈非編碼RNAs (lncRNAs)參與癌癥的多種生物學過程。然而,迄今為止,m5C甲基化的lncRNAs在胃癌(GC)中的研究很少。本研究中,作者發現RNA胞嘧啶- C(5)-甲基轉移酶(NSUN2) 在胃癌中上調,并且NSUN2的高表達與不良預后相關。lncRNA NR_033928發生NSUN2甲基化且在胃癌中表達上調。機制上,NR_033928作為IGF2BP3/HUR復合物的支架,提高谷氨酰胺酶(GLS)mRNA的穩定性,從而增加其表達。谷氨酰胺代謝物α-KG的積累促進NR_033928啟動子5-羥甲基胞嘧啶(hm5C)去甲基化,從而上調NR_033928的表達,形成正反饋環路。該研究于2023年8月發表在《Cell Death & Disease》,IF:9.0。
技術路線

主要研究結果
1. m5C調節因子上調NSUN2與胃癌患者不良預后相關
從癌癥基因組圖譜(TCGA)數據庫中檢測m5C在胃腺癌(STAD)項目中的表達。結果表明,與正常組織相比,m5C的writers(NOP2,NSUN2,NSUN3,NSUN4,NSUN5和NSUN6)和readers(ALYREF和YBX1)在胃癌樣本中上調。核心m5C甲基轉移酶NSUN2在GC中表達最高(圖1A)。Pearson相關分析表明,NSUN2的表達與大多數其他m5C調節因子的表達呈正相關(圖1B)。分析NSUN2在24對配對的胃癌和癌旁正常組織微陣列中的表達。結果顯示NSUN2在GC中顯著升高(圖1C)。在48對胃癌和匹配的正常胃組織中進行的qPCR分析驗證NSUN2在胃癌中的表達顯著較高(圖1D)。Western blot分析顯示NSUN2蛋白在胃癌組織中表達明顯升高(圖1E)。Kaplan-Meier Plotter結果顯示,NSUN2表達與胃癌患者的不良總生存期呈正相關(圖1F)。總之,這些結果表明NSUN2在胃癌中表達上調,并且與胃癌患者的不良診斷相關。
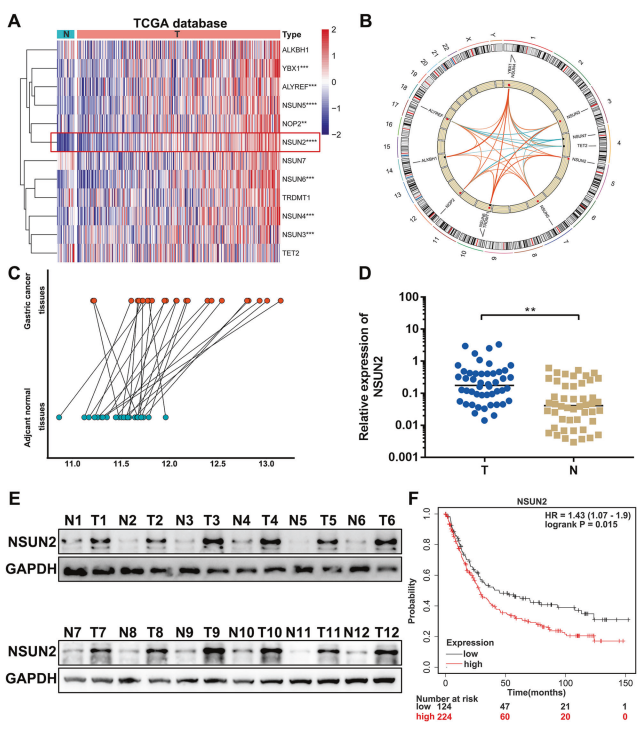
圖1. 在胃癌中NSUN2表達上調并與不良預后相關
2. NR_033928在GC中上調,且被鑒定為發生NSUN2甲基化的lncRNA
為繪制GC中lncRNA的m5C表觀遺傳修飾圖,作者對3對胃癌組織及其癌旁正常組織進行基于m5C抗體的RNA甲基化測序和二代測序。結果顯示,有254個表達失調和11107個不同程度甲基化的lncRNAs (圖2A,B)。進一步分析發現有10個上調且高甲基化的lncRNAs,1個上調且低甲基化的lncRNA,3個下調且高甲基化的lncRNAs,1個表達下調且低甲基化的lncRNA(圖2C)。在胃癌細胞中轉染NSUN2 siRNA,qPCR檢測NSUN2修飾的lncRNAs。結果顯示,在NSUN2沉默的細胞中,NR_033928的表達下調最明顯(圖2D)。qRT-PCR證實NR_033928在48例GC患者的GC組織中表達高于配對的正常組織(圖2E)。RNA熒光原位雜交(FISH)分析表明,與鄰近正常黏膜組織相比,GC組織具有顯著的NR_033928豐度(圖2G)。通過分析患者的臨床病理特征,發現NR_033928的表達水平與胃癌腫瘤大小和TNM分期呈正相關(表1)。檢測 NR_033928 在 GC 細胞系中的表達,結果顯示與正常人胃細胞系GES-1相比,NR_033928在所有檢測的胃癌細胞系(HGC-27, MKN28, MKN45, AGS和SNU1)中的表達均較高(圖2F)。根據上調的表達水平,選擇MKN45和AGS進行進一步研究。FISH顯示NR_033928主要定位于AGS和MKN45的細胞質中(圖2H)。CPC2和CPAT預測NR_033928的編碼概率極低(圖2I)。綜上所述,NR_033928被確定為胃癌中潛在的NSUN2修飾的致癌lncRNA。

圖2. NR_033928的鑒定與表征
3. NR_033928在體內和體外均有致癌作用
在GC細胞中進行功能增益和功能喪失試驗。將NR_033928 siRNA 和過表達載體分別轉染到MKN45和AGS細胞中。菌落形成實驗表明,敲除NR_033928 會減少MKN45細胞中的菌落數量,而過表達NR_033928則會促進AGS細胞中的菌落形成(圖3A,B)。同樣,EDU試驗表明,減少NR_033928的表達會削弱MKN45細胞的增殖能力,而增加NR_033928的表達則會增強AGS細胞的增殖能力(圖3C,D)。此外,流式細胞術檢測顯示,與對照組相比,轉染NR_033928 siRNA的MKN45細胞中凋亡細胞比例增加(圖3E)。而轉染NR_033928過表達載體的AGS細胞凋亡細胞數量減少(圖3F)。將轉染有NR_033928 shRNA和過表達載體的GC細胞皮下注射到5周大的Balb/c小鼠體內。異種移植腫瘤模型顯示,注射MKN45-sh-NR_033928小鼠的腫瘤重量和體積明顯小于對照組(圖 3G)。穩定轉染過表達NR_033928慢病毒的AGS細胞促進異種移植瘤的生長(圖 3H)。總之,NR_033928在體外和體內都促進GC的增殖,抑制細胞凋亡。
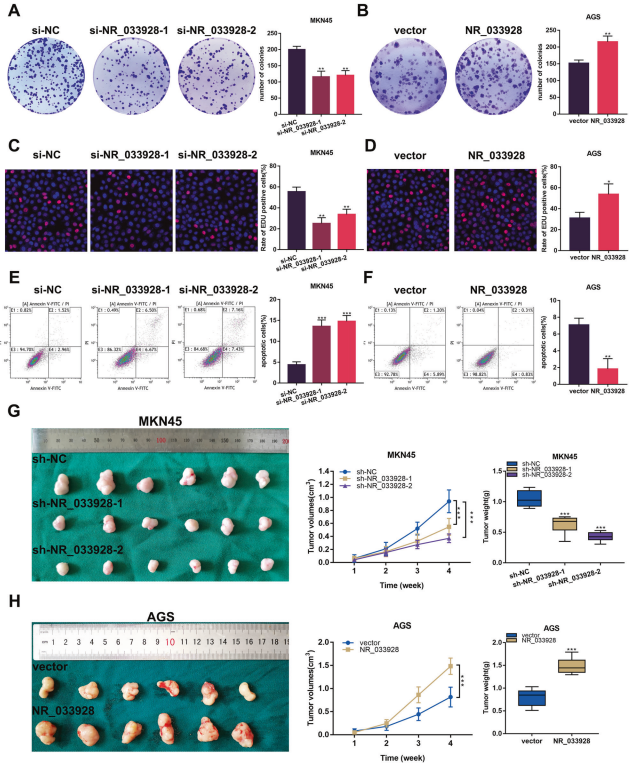
圖3. NR_033928促進細胞增殖,抑制細胞凋亡。
4. NSUN2通過維持NR_033928在GC中的穩定性而上調其表達
m5C RIP分析顯示,在GC細胞中同步轉染NSUN2 siRNA或過表達載體后,NR_033928的m5C修飾水平降低或升高(圖4A-D)。Sanger測序驗證特定的m5C甲基化位點C154(圖4E)。在敲低NSUN2后,這種甲基化被完全消除,堿基C被轉化為堿基T。通過Dactinomycin實驗評估NSUN2是否通過調節NR_033928的RNA穩定性來調節其表達。結果顯示,在轉染NSUN2 siRNA或過表達載體的細胞中,NR_033928的半衰期縮短或延長(圖4F-I)。綜上所述,結果表明NSUN2催化NR_033928的m5C修飾,并通過增強RNA穩定性來上調其表達。
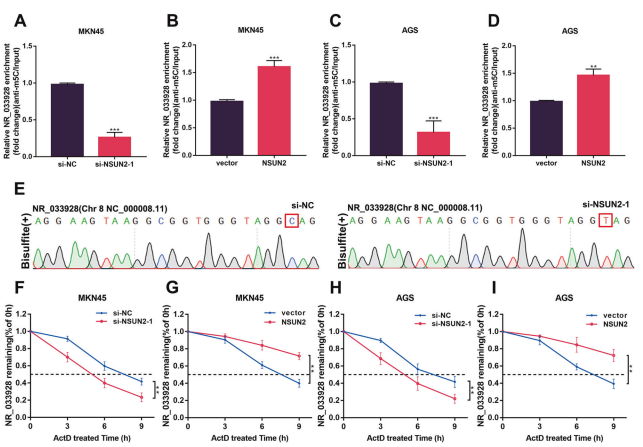
圖4. NSUN2介導NR_033928 m5C甲基化并調控其表達。
5. NR_033928通過GLS介導的谷氨酰胺代謝調控胃癌的增殖和凋亡
在NR_033928缺陷和野生型細胞中進行RNA-seq(圖5A)。通過KEGG分析發現NR_033928的表達與谷氨酰胺代謝通路呈正相關,而谷氨酰胺代謝通路與癌癥進展密切相關(圖5B)。EDU實驗表明NR_033928過表達細胞在正常培養基和谷氨酰胺剝奪培養基中增殖能力增強或減弱(圖5C)。細胞凋亡實驗顯示,正常培養基中NR_033928過表達的細胞凋亡比例降低,而在去除谷氨酰胺的培養基中,凋亡細胞的比例恢復(圖5E)。在所有差異表達基因中,GLS是參與谷氨酰胺分解的關鍵基因。在GC細胞中,過表達NR_033928上調谷氨酸和α-KG的含量,而共轉染sh-GLS則下調它們的含量(圖5D, F)。EDU實驗顯示,在AGS細胞中,過表達NR_033928增加EDU陽性細胞的比例,在NR_033928過表達載體和sh-GLS共轉染后,EDU陽性細胞的比例下降(圖5G)。細胞凋亡實驗顯示,在AGS細胞中過表達NR_033928降低凋亡細胞的比例,而在AGS細胞中共轉染NR_033928過表達載體和sh-GLS挽救凋亡細胞的比例(圖5H)。在AGS細胞中應用GLS抑制劑Telaglenastat(CB-839)。EDU分析顯示CB-839降低AGS細胞中轉染NR_033928過表達載體的細胞的增殖活性(圖5I)。凋亡分析表明,CB-839促進AGS細胞中轉染NR_033928過表達載體的細胞的凋亡(圖5J)。這些結果表明,NR_033928通過GLS介導的谷氨酰胺代謝在胃癌中發揮作用。

圖5. NR_033928通過GLS介導的谷氨酰胺代謝促進GC進展
6. NR_033928與IGF2BP3/HUR復合物相互作用并促進其形成
在胃癌細胞中分別進行3次RNA pull-down和質譜分析。在所有的候選蛋白中,IGF2BP3和HUR存在于質譜結果和預測的GLS結合蛋白結果中 (圖6A-C)。使用IGF2BP3和HUR抗體進行RNA免疫沉淀分析。結果顯示,與IgG組相比,IGF2BP3或HUR抗體組中NR_033928的表達顯著較高(圖6D)。RNA pull-down分析證實,生物素標記的NR_033928可以沉淀出重組IGF2BP3和HUR(圖6E)。提出假設NR_033928作為IGF2BP3/ HUR復合物的支架來促進GLS mRNA在胃癌中的穩定性。免疫共沉淀實驗表明,IGF2BP3和HUR在GC中相互作用(圖6F, G)。siRNA沉默NR_033928,減少IGF2BP3或HUR抗體沉淀的HUR或IGF2BP3的蛋白量(圖6H, I)。為進一步闡明NR_033928如何招募IGF2BP3和HUR,設計NR_033928缺失突變體。RNA pull-down分析顯示,含有1387-1926nt的NR_033928 #1片段與IGF2BP3相互作用,含有1387-1926nt、50-1014nt、1 - 1374nt的#1、#4、#5片段與HUR相互作用(圖6J)。此外,根據IGF2BP3和HUR的內在蛋白結構域構建一系列flag標記的截短體。RIP實驗表明,IGF2BP3結構域(198-344aa)和HUR結構域(1-99aa)與NR_033928特異性相互作用(圖6K, L)。總之,NR_033928與IGF2BP3/HUR復合體相互作用并促進其形成。
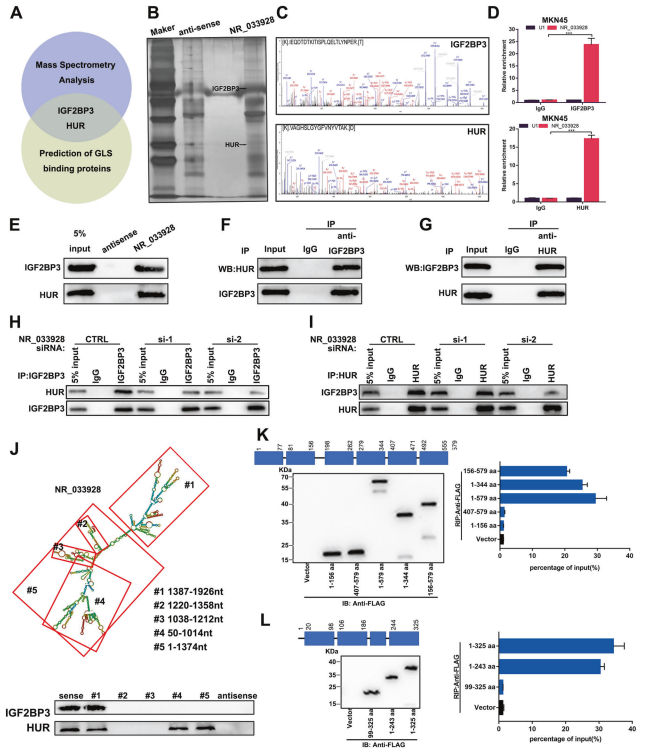
圖6. NR_033928直接結合IGF2BP3/HUR復合物
7. NR_033928促進IGF2BP3/HUR復合物與GLS mRNA的相互作用
RIP實驗表明HUR和IGF2BP3與GLS結合(圖7A,E)。Western blot分析證實,構建IGF2BP3和HUR的過表達質粒和siRNA,并在GC細胞中轉染(圖7B,F)。Dactinomycin實驗表明,過表達IGF2BP3使GLS的半衰期延長,共轉染HUR siRNA使GLS的半衰期縮短(圖7C)。在使用HUR過表達載體和IGF2BP3 siRNA進行的Dactinomycin檢測中,也檢測到類似的結果(圖7G)。這些結果表明,IGF2BP3和HUR協同增強GLS的穩定性。RIP實驗顯示,沉默NR_033928或過表達NR_033928降低或增加IGF2BP3和HUR結合的GLS的數量(圖7D,H)。Dactinomycin實驗顯示,si-IGF2BP3或si-HUR縮短GLS的半衰期,與NR_033928過表達載體共轉染延長GLS的半衰期(圖7I,K)。qRT-PCR分析表明,抑制IGF2BP3表達降低通過過表達NR_033928增加的GLS表達(圖7J)。同樣,抑制HUR表達降低GLS的表達,而過表達NR_033928增加GLS的表達(圖7L)。因此,NR_033928通過作為IGF2BP3/HUR復合物和GLS的支架調節GLS的穩定性,從而調節GLS的表達。

圖7. NR_033928在IGF2BP3/HUR和GLS之間起支架作用
8. α-KG促進NR_033928啟動子去甲基化從而增加NR_033928的表達
NR_033928在谷氨酰胺缺失培養基中表達顯著降低(圖8A)。沉默GLS降低NR_033928的表達(圖8B)。qRT-PCR分析表明,添加外源性合成的α-KG (DM-α-KG)而不是谷氨酰胺的其他代謝物增加NR_033928的表達(圖8C)。α-KG異常積累作為DNA去甲基化酶(TETs)和組蛋白去甲基化酶(JMJDs)的輔因子在基因組表觀遺傳調控中發揮重要作用(圖8D)。通過GSK-J4抑制組蛋白去甲基化酶(JMJDs)并沒有改變NR_033928的表達(圖8E)。Tet家族(Tet1 Tet2和Tet3)的聯合沉默顯著降低NR_033928的表達(圖8F)。hMeDIP實驗表明,添加外源性DM-αKG增加NR_033928啟動子hm5C的水平,在谷氨酰胺去除培養基培養的細胞中轉染sh-Tet1-3降低hm5C的水平(圖8G,H)。總之,α-KG以TETs依賴的DNA去甲基化方式促進NR_033928的表達。
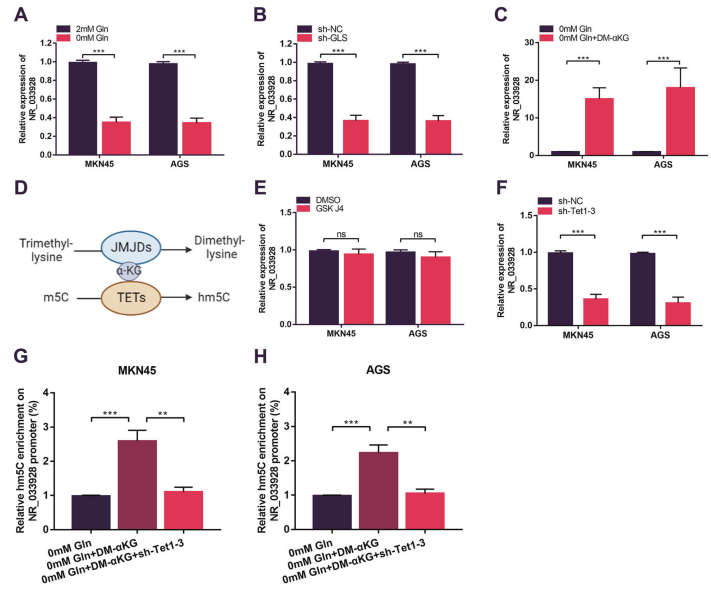
圖8. α-KG通過促進NR_03928啟動子去甲基化上調NR_033928的表達
9. NR_033928在GC中的臨床意義
對隨訪數據的Kaplan-Meier分析表明,NR_033928的表達與患者的總生存期呈負相關(圖9A)。此外,在線Kaplan-Meier模型結果顯示,GLS高表達的患者總生存期低于GLS低表達的患者(圖9B)。在異種移植瘤模型中,第4周處死所有小鼠,并將腫瘤用于免疫組織化學分析(圖9C)。GLS免疫組織化學染色顯示,敲低NR_033928后,GLS的表達明顯降低。此外,對增殖標志物Ki67和凋亡標志物c-caspase3的分析表明,沉默NR_033928降低Ki-67的表達,增強c-caspase3的表達。因此,NR_033928可作為胃癌的潛在預后標志物,沉默NR_033928可抑制胃癌細胞增殖并促進其凋亡。

圖9. NR_033928作為胃癌的預后和治療性生物標志物
結論
綜上所述,本研究發現一種NSUN2甲基化的lncRNA,NR_033928,在GC細胞和組織中高表達。NR_033928通過GLS 介導的谷氨酰胺代謝促進GC的進展,可能是一個潛在的預后和治療靶點。
實驗方法
細胞培養,m5C MeRIP測序和MeRIP- PCR,lncRNA表達譜和mRNA二代測序,寡核苷酸、慢病毒和質粒轉染,RNA提取和PCR,FISH,WB,RNA半衰期檢測,菌落形成實驗,EDU染色,細胞凋亡檢測,谷氨酸和α-酮戊二酸檢測,羥甲基化DNA免疫沉淀,裸鼠實驗,免疫組化,RIP,Co-IP,RNA pull-down和質譜分析,sanger測序
參考文獻
Fang L, Huang H, Lv J, Chen Z, Lu C, Jiang T, Xu P, Li Y, Wang S, Li B, Li Z, Wang W, Xu Z. m5C-methylated lncRNA NR_033928 promotes gastric cancer proliferation by stabilizing GLS mRNA to promote glutamine metabolism reprogramming. Cell Death Dis. 2023 Aug 15;14(8):520. doi: 10.1038/s41419-023-06049-8. PMID: 37582794; PMCID: PMC10427642.