






基于人類PBMC scRNA-seq的衰老時鐘揭示了 核糖體與炎癥平衡作為單細胞衰老和超長壽命的標志
量化老年率對于評估與年齡相關的衰老和死亡率很重要。最近生成了一個七位超級百歲老人(SCs)的血液單細胞RNA測序數據集。在這里,作者生成一個參考28個樣本的年齡隊列,以計算單細胞水平的老化時鐘,并確定SCs的生物年齡。作者的時鐘模型將SCs的血液生物學年齡定在80.43歲到102.67歲之間。與模型預期的衰老軌跡相比,SCs表現為原始CD8+T細胞增加,細胞毒性CD8+T細胞減少,記憶CD4+T細胞和巨核細胞減少。作為單細胞水平上最顯著的分子標志,SCs含有更多的高核糖體水平的細胞和細胞類型,根據貝葉斯網絡推斷,這與SCs的低炎癥狀態和緩慢衰老有關。通過抑制單核細胞中的核糖體活性或翻譯過程,驗證了單細胞衰老時鐘揭示的翻譯與炎癥平衡的關系。
該研究于2023年6月發表在《Science advances》,IF:14.957。
技術路線
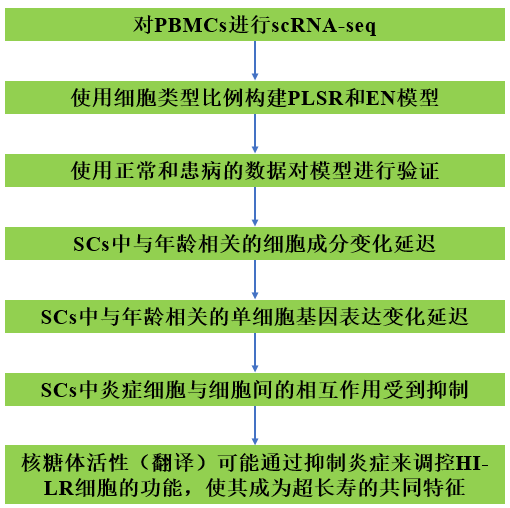
結果
1、scRNA-seq揭示了衰老隊列中的細胞類型組成
為了構建基于單細胞轉錄組的衰老速率預測模型,作者從28歲到77歲的17個個體(其中6個男性和11個女性,年齡分布相似)收集了PBMCs(外周血單核細胞),作者將其稱為上海東方醫院自然衰老隊列(簡稱SE隊列),并使用10X Genomics scRNAseq對其進行測序。對于每個個體,平均測序和分析了8545個細胞,每個細胞平均有110,364個reads(圖1A)。作者將這些數據與已發表的PBMC scRNAseq數據集一起進行進一步分析。這些數據集包括兩個中國隊列,中國年輕隊列(CYCT)和武漢隊列(WHCT),以及兩個日本隊列,日本老年隊列(JOCT)和超百歲人士隊列(SCs)。每個隊列包含五個個體。由于質量不佳(線粒體基因表達超過10%或檢測到的基因數小于200),作者移除了25,111個細胞,總共保留了131,972個單細胞的SE數據集,以及23,796個細胞的CYCT數據集,45,923個細胞的WHCT數據集,19,252個細胞的JOCT數據集和38,417個細胞的SC數據集。然后,作者消除了SE數據集和其他獨立隊列之間的批次效應,并根據經典細胞系標記物和簇特異性標記基因的表達,識別了紅細胞(RBCs)、巨核細胞(Mega)和六個主要的免疫細胞系譜,包括CD4+ T細胞(TC)、CD8+ T細胞、自然殺傷細胞(NK)、單核細胞(MC)和樹突狀細胞(DC),并通過UMAP進行可視化(圖1B)。
2、免疫細胞亞型重新聚類進行細分
為了分離免疫細胞的亞型,作者分別對NK和TC系譜的細胞、BC系譜的細胞以及MC和DC系譜的細胞進行了重新聚類。利用每個簇中最顯著上調的基因,作者總共確定了29個細胞亞型(圖1B)。這些細胞亞型隨后被用于下游分析。

圖 1. 衰老過程中人血外周血單核細胞 (PBMC) 的單細胞 RNA 測序 (scRNA-seq) 圖譜
3、基于細胞類型比例的時鐘模型顯示,SCs比實際年齡年輕得多
隨著年齡的增長,各種細胞類型的比例發生變化(圖2A)。因此,作者測試使用scRNA-seq數據中細胞類型比例來構建一個"時鐘"來預測年齡。作者首先訓練了一個偏最小二乘回歸(PLSR)線性模型,并使用留一法交叉驗證(LOOCV)來評估模型的準確性。作者計算了變量在投影中的重要性(VIP)來對每個變量對PLSR模型的整體貢獻進行排序,VIP得分>1的變量被認為是高解釋變量。這些變量包括NK-GZMH、CD8-CTL、CD8-Naive、CD4-Tm、CD4-Naive、Naive-B、NK-FCER1G、CD14-MC和Memory-B-CRIP1(圖2B)。為了擴展訓練數據的年齡范圍,作者使用TOSICA,進一步映射了由廣東醫科大學產生的包含非常老(72至100歲)樣本(GM)的公開PBMC數據集的細胞類型,并訓練了一個PLSR模型。作者發現它們在細胞類型組成或scRNA-seq時鐘預測的cAgeDiff中并沒有顯著差異。因此,作者將這兩組樣本都包含在擴展的訓練樣本中。基于這個擴展數據集(SE + GM),作者發現僅基于前7個細胞類型的三個成分的PLSR模型產生了一個預測年齡與實際年齡之間相關系數(PCC)為0.88,平均絕對誤差(MAD)為8.36的模型(圖2C)。作者還使用細胞類型比例來訓練一個彈性網絡(EN)回歸線性模型,并使用權重來對每個變量對EN模型的整體貢獻進行排序。基于前7個細胞類型的EN模型具有與PLSR模型類似的準確預測能力,在SE + GM隊列中預測年齡與實際年齡之間的PCC和MAD分別為0.89和7.86,而PLSR模型的準確性在訓練和獨立隊列之間更為相似,表明稍微具有更好的泛化能力。PLSR模型和EN模型之間前7個細胞類型有5個共同的,暗示了它們在通過血液細胞類型組成進行年齡預測中的重要性。
為了進一步驗證作者的模型,作者將其應用于三個獨立健康隊列的血液scRNA-seq數據上。其中包括五個CYCT個體(年齡范圍為29至58歲)、五個WHCT個體(年齡范圍為37至71歲)和五個JOCT個體(年齡范圍為50至80歲,年齡按十年為單位精確,因此根據定義,預期的誤差超過5年)。在這三個隊列中,作者的七種細胞類型的PLSR模型在按年齡排序三個隊列中的MAD分別為10.06、10.91和10.16,PCC分別為0.43、0.69和0.28。當繪制預測年齡與實際年齡之間的差異(AgeDiff)時,注意到在PLSR和EN模型中老年人中存在輕微的系統性低估(negative AgeDiff),而在年輕人中存在高估(positive AgeDiff)。因此,作者使用Loess模型進行了系統性偏差的校正,得到了校正后的AgeDiff(cAgeDiff)。在這種校正后,獨立對照隊列中與實際年齡的MADs為7.95、8.19和8.62,PCC為0.64、0.85和0.57。值得注意的是,年齡校正不會改變模型與實際年齡的MAD,但會增強PCC。由于作者的單細胞衰老時鐘與年齡呈線性關系,作者希望它具有線性可擴展性,可以外推到訓練數據中未包含的年齡譜的任何一端。根據作者的時鐘模型預測,比最小的訓練樣本小28歲以上,TOSICA映射的臍帶血樣本(Cord)的年齡為-10.10至-5.42歲(圖2D)。而在年齡范圍的另一端,比最年長的訓練樣本年大20歲的SCs被預測為80.43至102.67歲,平均比他們的實際年齡年輕18.64歲。經過年齡校正的細胞類型EN模型將所有SCs預測為低于110歲,平均年齡為95.23歲,將臍帶血(Cord)預測為7.12至12.68歲。
為了進一步測試時鐘性能,將年齡時鐘應用于其他獨立的疾病數據集,包括COVID-19數據集和系統性紅斑狼瘡(SLE)數據集。基于細胞類型組成的PLSR模型顯示,處于疾病狀態的個體的平均生物學年齡高于年齡匹配的對照組,并且隨著疾病嚴重程度的增加而增加(圖2,E至G)。也許由于樣本規模較小,只有嚴重癥狀的COVID-19患者明顯年齡較大(圖2E;t檢驗,P = 0.040),而SLE癥狀得到控制和病情加重的患者年齡較明顯大于對照組(圖2G;t檢驗,P = 8.77 × 10?4和4.20 × 10?3)。由EN模型預測的cAgeDiff顯示出疾病與對照組之間一致顯著的增加。這些結果表明,基于細胞類型組成的PLSR和EN時鐘模型都能夠識別這些疾病患者免疫相關生物年齡的增加。
與模型預測的SCs的“緩慢老化”一致(圖2D),在PLSR和EN模型中使用的七個細胞類型中,兩個CD8+T細胞亞群之間存在高度顯著的負相關,即CD8+ CTL和CD8+ 初始 T細胞,在SE + GM和SC隊列中,它們分別隨年齡顯著增加和減少,但SCs在很大程度上與70至100歲的SE + GMs個體重疊(圖2H)。這表明,在血液細胞群體水平上,CD8+ 初始T細胞向CD8+ CTL的極化是衰老的標志。

圖 2. 基于單細胞 RNA 測序 (scRNA-seq) 的單細胞成分衰老時鐘和超級百歲老人 (SCs) 的年齡延遲。
4、SCs中與年齡相關的細胞成分變化被推遲
作者利用模型預測與SCs實際狀態之間的細胞和分子偏差,來解析其極端長壽相關的特征(圖3A)。作者通過PLSR時鐘模型反向計算了110歲時的細胞類型組成。另外,基于PLSR和EN時鐘模型中的前7個細胞類型(圖2E),作者使用線性回歸模型(LR)定義了與年齡顯著正相關或負相關的細胞類型(具有RCC與年齡斜率,P < 0.05)。然后,根據PLSR模型或LR模型計算了110歲時的預期細胞比例,如果它們與SCs中的實際比例相比顯著偏高或偏低(z分數,P < 0.05),則將這些細胞類型視為“SC延遲衰老細胞類型”或“SC延遲年齡下降細胞類型”(圖3A)。作者發現,與PLSR模型相比,SCs在CD8+ 初始T細胞、初始B細胞、記憶B細胞和NK-GZMK方面顯示出明顯的延遲性年齡相關性下降(z分數,P < 0.05),在CD4+ Tm方面顯示出較小的延遲衰老相關增加(z分數,P < 0.1),在CD8+ 初始T細胞方面顯示出延遲衰老相關減少,并在CD4+ Tm和初始B細胞方面顯示出延遲衰老相關增加,與LR模型相比。當繪制年齡下調的CD8-初始細胞和年齡上調的CD4 Tm細胞的比例隨年齡變化時,在28到100歲的28個樣本中,SCs與線性趨勢線呈現明顯的延遲偏差。
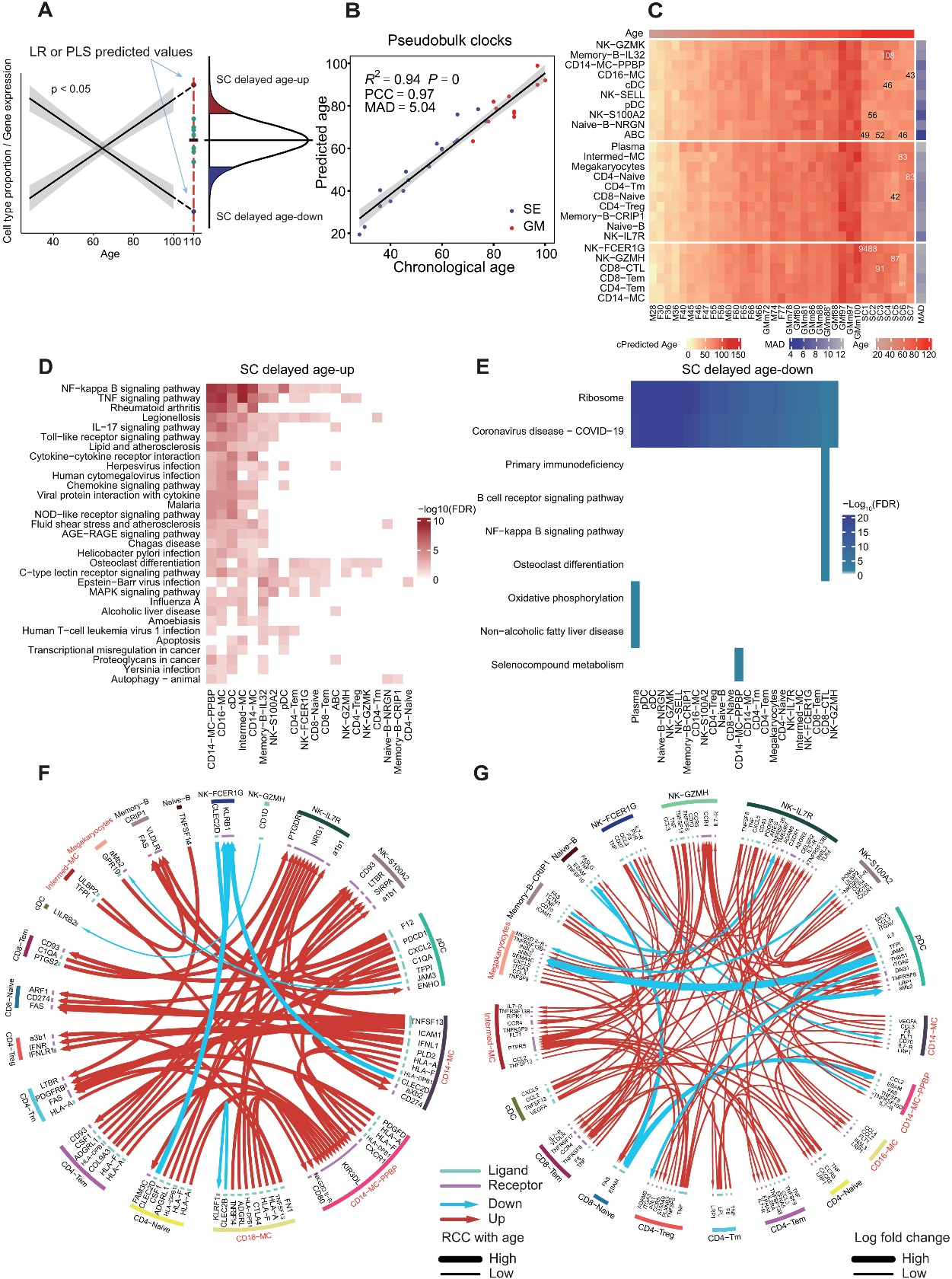
圖 3. 超級百歲老人 (SCs) 中與衰老相關的細胞類型比例和基因表達變化延遲。
5、SCs中與年齡相關的單細胞基因表達變化延遲
作者使用每個個體所有細胞的基因表達之和來訓練PLSR模型,作為偽批量轉錄組時鐘模型,該模型對28個訓練樣本的預測與實際年齡的MAD為5.04,PCC為0.97(圖3B),并將SCs的平均年齡預測為82.49歲。作者還使用每個細胞類型每個個體所有細胞基因表達的總和,分別訓練了各細胞類型的偽批量轉錄組時鐘模型,MAD在4.80年(ABC細胞)到9.60年(漿細胞)之間(圖3C)。與基于細胞類型比例的時鐘模型相比,這些時鐘模型更加顯著地將SCs的中位轉錄組年齡預測為51.62歲到63.19歲(年齡校正前),年齡校正后為65.01歲到75.83歲(圖3C)。接著,基于這些PLSR時鐘模型,或者根據貢獻度VIP > 1的基因(表S1)定義為與年齡相關的基因,使用LR模型在每個細胞類型中預測它們在110歲時的表達水平,然后類似于上面描述的細胞類型分析,定義了“SC延遲年齡增加基因”(與年齡上調,并且在SCs中延遲)和“SC延遲年齡下調基因”(與年齡下調,并且在SCs中延遲)(圖3A)。使用LR模型,該模型不顯示對年齡的系統性偏差,作者發現2749個年齡增加基因和495個年齡下調基因,并且1023個年齡增加基因和149個年齡下調基因在SCs中有顯著的延遲表達。轉錄因子(TF)靶標富集分析顯示,年齡增加和年齡下調基因都富集了FOXR2、ZNF318、PSMB5、PER1、ZZZ3和NERF的靶。其中,FOXR2、ZNF318、PSMB5和ZZZ3的靶標在SC延遲年齡下調基因中富集,核因子κB(NFκB)的靶標在SC延遲年齡增加基因中富集。KEGG通路富集分析顯示,年齡增加基因富集了NFκB/TNF/TLR/NOD/FoxO信號通路、病毒感染、凋亡、T細胞受體信號通路、細胞因子和PD-1通路,其中,NFκB/TNF/TLR/NOD/FoxO信號通路、病毒感染、凋亡、細胞因子和絲裂原活化蛋白激酶(MAPK)通路在SCs中有顯著的延遲(圖3D),而年齡下調基因主要富集于核糖體基因,這些基因在SCs的大多數細胞類型中顯著延遲表達(圖3E)。
與PLSR模型預期值相比,SCs中有111個顯著延遲表達的年齡上調基因,與MAPK信號通路、雷帕霉素(mTOR)信號通路、NFκB信號通路和病毒感染有關。另外,有1261個年齡下調基因在SCs中顯著延遲表達。再次,大部分細胞類型中富集了與核糖體和翻譯相關的基因。而在其他隊列中,SEs中鑒定出的年齡下調核糖體基因也隨著年齡的增加而減少。
6、SCs中炎癥細胞與細胞間的相互作用受到抑制
通過使用細胞間通訊數據庫,作者計算了每對細胞類型之間的配體-受體對數量,利用CellphoneDB進行分析。作者發現,在不同的細胞類型中,許多與衰老相關的受體-配體對在衰老過程中上調(PCC,P < 0.05)。這些受體-配體對主要與炎癥、細胞因子和抗原呈遞相關(圖3F)。類似于延遲上調和下調的細胞類型和基因,作者定義了在SCs中延遲上調和下調的配體-受體對。與大多數細胞類型相比,與年齡相關的下調的aMb2整合素信號配體-受體對在SCs中普遍延遲,而與炎癥相關的上調的TNF信號配體-受體對也被延遲,特別是在pDC中(圖3G)。
7、核糖體/翻譯在單細胞水平上對抗炎癥
根據上述結果,作者想知道核糖體水平的年齡相關下降和炎癥水平的年齡相關增加是否是單個細胞老化的兩個相關事件。為了定量評估每個細胞的炎癥狀態,作者根據炎癥相關基因列表計算了每個細胞的炎癥分數。核糖體基因的中位表達水平被用來指示每個細胞中核糖體的水平。在SE和SC隊列中,對于所有細胞類型的所有單個細胞,核糖體水平與炎癥分數之間存在顯著的負相關(圖4A)。此外,在不同的細胞類型之間,平均核糖體水平和平均炎癥分數之間也存在顯著的負相關(圖4B)。根據核糖體水平與炎癥分數的分布,細胞類型可以分為高炎癥和低核糖體(HI-LR)細胞類型,包括Intermed-MC、CD14-MC、CD16-MC、CD14-MC-PPBP和巨核細胞,以及低炎癥和高核糖體(LI-HR)細胞類型,即其他細胞類型(圖4B)。在除一種細胞類型外,所有核糖體基因的表達水平都隨著年齡的增長而在SE中降低,在HI-LR細胞類型中,SCs的水平與年輕的SEs相似(圖4C)。一致地,基于每個細胞類型內的核糖體和炎癥基因表達的主成分分析(PCA)顯示,對于大多數細胞類型(例如CD4-Treg和CD14-MC),單個細胞分別落入高和低核糖體表達的兩個主要細胞狀態。與核糖體表達水平一致,大多數細胞類型中處于高核糖體狀態的細胞比例隨年齡減少(圖4D)。值得注意的是,在HI-LR細胞類型中,SCs始終比SE擁有更多處于高核糖體狀態的細胞(圖4D)。這表明,在血液中,高炎癥低核糖體表達,或者更準確地說,在高核糖體狀態下的細胞更多,是超長壽在單個細胞水平上的共同特征。
作者還在COVID-19和SLE隊列中驗證了核糖體表達和炎癥分數的變化。與核糖體表達和炎癥之間的平衡一致,在COVID-19和SLE的疾病進展中,所有細胞類型和HI-LR細胞中炎癥分數持續增加,核糖體表達水平持續下降,而在SLE中的變化比COVID-19更顯著。
此外,作者發現大多數細胞類型內處于高核糖體狀態與低核糖體狀態的細胞之間的細胞間通信顯著不同(圖4E)。在低核糖體狀態下,前100個配體-受體對主要是在CD8-CTL和其他細胞類型之間,包括MHCII、S100A8/A9TLR4、CCL5-CCR5和TNF相關的配對,而在高核糖體狀態下這些配對顯著降低(t檢驗,P < 0.01;圖4E)。在高核糖體狀態與低核糖體狀態中,最常見的下調配體-受體對是核糖體蛋白S19(RPS19)與其膜受體C5AR1,據報道,該相互作用能抑制免疫反應。
最后,為了了解SE + GM數據中核糖體水平、炎癥水平、年齡和衰老速率之間的因果關系,作者分別基于HI-LR和LI-HR細胞類型中的所有單個細胞,在衰老速率參考SE + GM數據中推斷了一個貝葉斯網絡(BN)。這里,衰老速率通過年齡校正的AgeDiff來衡量,AgeDiff是基于細胞組成PLSR模型預測年齡與實際年齡之間的差異,并經過年齡校正。BN模型推斷出AgeDiff取決于HI-LR細胞類型中的核糖體水平和炎癥水平,而核糖體水平反過來又取決于年齡。模型還推斷出核糖體水平的降低會導致HI-LR細胞中炎癥水平的增加。然而,在LI-HR細胞類型中,年齡降低會降低炎癥分數,并且炎癥分數不會影響衰老速率(圖4F)。這表明核糖體豐度,通常是翻譯和生長速率的代表,可能抑制HI-LR細胞的炎性細胞因子分泌;換句話說,高水平的炎性細胞因子表達可能是HI-LR細胞類型生長停滯或基礎低核糖體活性不足的結果,而干細胞可能受益于高細胞核糖體/翻譯/生長速率以對抗衰老引起的慢性炎癥。相比之下,基礎水平的炎性細胞因子可能是LI-HR細胞正常生長的結果(圖4F)。對于每種細胞類型推斷的BN模型中可以一致地觀察到類似的關系,根據這些BN中的相互作用,HI-LR細胞類型會自動聚集在一起,并通過無監督層次聚類與其他細胞類型分離開來(圖4G)。
最后,為了確認HI-LR細胞中核糖體活性(翻譯)對炎癥的抑制作用,作者在人類血液原代單核細胞和THP-1單核細胞系(代表HI-LR細胞類型,并且在作者先前的研究中發現其分泌衰老相關的細胞因子)中,使用翻譯抑制劑環己基甲酰胺(CHX)進行時間限制的翻譯抑制。CHX能與60S核糖體亞單位結合,抑制翻譯延伸。如BN模型所預期的那樣,在原代單核細胞中,抑制翻譯引起了常見炎性細胞因子白細胞介素-6(IL-6)和IL-8的顯著上調,并且這種上調呈時間依賴性(圖4H)。類似的結果也在THP-1細胞中觀察到。血液單細胞衰老和 SC 延遲衰老標志的核糖體和炎癥平衡的示意圖(圖4I)。
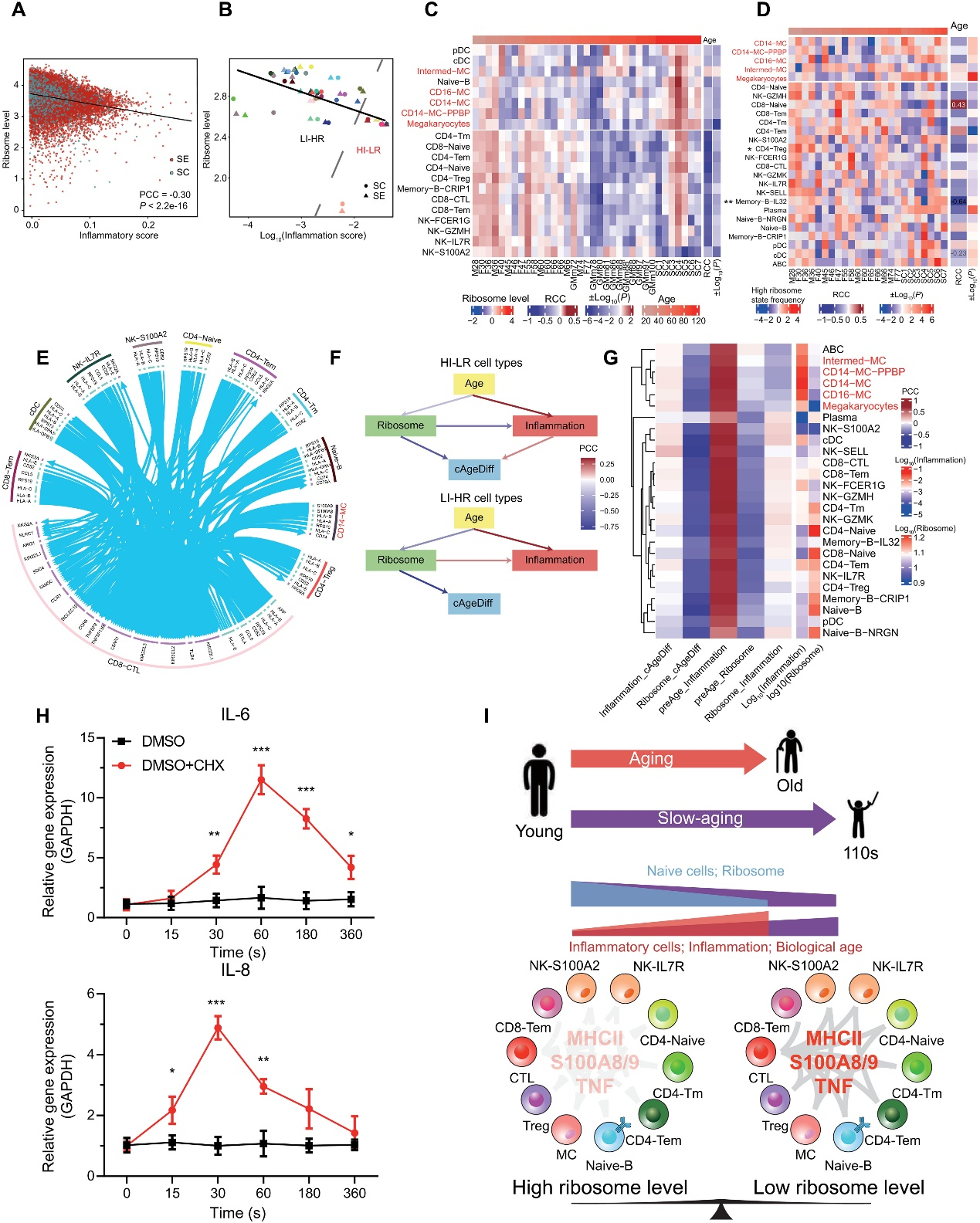
圖 4. 衰老過程中核糖體與單細胞和細胞類型炎癥之間的關系
實驗方法
scRNA-seq、校正批次效應、細胞亞型注釋、構建時鐘模型、飽和度分析、TOSICA、GO terms、TF 富集分析、炎癥評分、細胞間通訊分析、AgeDiff校正、BN推斷、翻譯抑制測定。
參考文獻
Hongming Zhu et al. ,Human PBMC scRNA-seq–based aging clocks reveal ribosome to inflammation balance as a single-cell aging hallmark and super longevity.Sci. Adv.9,eabq7599(2023).DOI:10.1126/sciadv.abq7599