






姜黃素激活ROS/KEAP1/NRF2/miR-34a/b/c級聯(lián)抑制結(jié)直腸癌轉(zhuǎn)移
姜黃素是從姜黃根中分離出來的一種天然植物化學(xué)物質(zhì),是預(yù)防和治療結(jié)直腸癌/CRC的候選藥物。作者使用遺傳方法來確定p53/miR-34通路作為姜黃素作用的中介作用。將三種p53、miR-34a和/或miR-34b/c基因缺失的等基因CRC細(xì)胞系暴露于姜黃素中并進(jìn)行細(xì)胞生物學(xué)分析。通過siRNA介導(dǎo)的NRF2抑制和異位表達(dá),并對其靶基因進(jìn)行Western blot、qPCR和qChIP分析。將結(jié)直腸癌細(xì)胞靜脈注射到NOD/SCID小鼠體內(nèi),通過縱向無創(chuàng)成像確定肺轉(zhuǎn)移的形成。在結(jié)直腸癌細(xì)胞中,姜黃素誘導(dǎo)細(xì)胞凋亡和衰老,并以p53不依賴的方式抑制細(xì)胞的遷移和侵襲。姜黃素通過誘導(dǎo)ROS激活KEAP1/NRF2/ARE通路。此外,姜黃素以ROS/ NRF2依賴和p53獨立的方式誘導(dǎo)miR-34a和miR-34b/c的表達(dá)。NRF2通過占據(jù)miR-34a和miR-34b/c啟動子區(qū)域的多個ARE基序直接誘導(dǎo)miR-34a和miR-34b/c。姜黃素逆轉(zhuǎn)了IL-6和缺氧誘導(dǎo)的miR-34a和miR-34b/c的抑制。缺失miR-34a和miR-34b/c可顯著降低姜黃素誘導(dǎo)的細(xì)胞凋亡和衰老,阻止姜黃素或異位NRF2對遷移和侵襲的抑制。在結(jié)直腸癌細(xì)胞中,姜黃素誘導(dǎo)MET并以miR34a依賴的方式阻止小鼠肺轉(zhuǎn)移的形成。此外,作者發(fā)現(xiàn)姜黃素可以增強5-FU對缺乏p53和miR-34a/b/c的CRC細(xì)胞的治療作用。KEAP1/NRF2/miR-34a/b/c軸的激活可介導(dǎo)姜黃素的腫瘤抑制活性,為激活腫瘤中miR-34基因以達(dá)到治療目的提供了一種新的途徑。本文于2023年5月發(fā)表在《Cell Death & Differentiation》IF:12.4期刊。
技術(shù)路線

主要實驗結(jié)果
1、姜黃素對結(jié)直腸癌細(xì)胞的p53非依賴性作用
姜黃素處理HCT116細(xì)胞后,p53蛋白的數(shù)量增加(圖1A)。為了確定p53與姜黃素對CRC細(xì)胞影響的相關(guān)性,作者用野生型p53和純合缺失p53的等基因HCT116細(xì)胞與姜黃素濃度增加一起處理48小時。存在p53的細(xì)胞顯示IC50為21.49 μM,而缺乏p53的細(xì)胞的IC50為18.03 μM (圖1B)。在p53存在或p53缺乏的RKO和SW48 CRC細(xì)胞系中觀察到類似的效果。姜黃素濃度為15 μM,略低于p53缺陷細(xì)胞的IC50值,用于后續(xù)實驗。通過阻抗測量(圖1C)證明了暴露于姜黃素強烈降低了p53缺陷和p53存在的HCT116細(xì)胞的增殖,并且在最終時間點證實了細(xì)胞數(shù)量的變化(圖1D)。姜黃素在結(jié)直腸癌細(xì)胞系RKO和SW48中也觀察到類似的p53非依賴性作用。姜黃素誘導(dǎo)的細(xì)胞活力和增殖速度的下降在p53缺陷細(xì)胞中更為明顯。因此,盡管p53在姜黃素治療后會積累,姜黃素對CRC細(xì)胞的這些作用是獨立于p53的。
接下來,作者分析了哪些過程可能是姜黃素介導(dǎo)的增殖抑制的基礎(chǔ)。姜黃素導(dǎo)致細(xì)胞中sub-G1 DNA含量增加,表明凋亡增加,與p53狀態(tài)無關(guān)(圖1E)。姜黃素處理后24小時,p53缺陷細(xì)胞表現(xiàn)出更多的G2/ m -阻滯,而WT p53細(xì)胞的G0/G1積累更多。膜聯(lián)蛋白V/ PI檢測顯示,與p53正常細(xì)胞相比,p53缺陷細(xì)胞的凋亡增加更為明顯(圖1F和S1G)。這些結(jié)果通過檢測被切割的parp、被切割的Caspase-3、Bcl-2和Bax蛋白得到了證實(圖1G、S1H和S1I):在p53缺失的細(xì)胞中,24小時檢測到Bax和被切割的Caspase 3的增加,而p53成熟的細(xì)胞顯示這些蛋白的誘導(dǎo)延遲了48小時。此外,通過檢測β-gal pH 6,姜黃素在p53精通和p53缺乏的HCT116細(xì)胞中誘導(dǎo)HCT116細(xì)胞衰老的程度相似(圖1H)。最后,姜黃素抑制了p53精通和p53缺乏的HCT116細(xì)胞的遷移和侵襲,后者顯示出更高的基礎(chǔ)遷移和侵襲水平(圖1I, J)。綜上所述,姜黃素抑制了細(xì)胞活力、增殖、遷移和侵襲,而以不依賴p53的方式誘導(dǎo)結(jié)直腸癌細(xì)胞的凋亡和衰老。

圖1姜黃素對結(jié)直腸癌細(xì)胞的p53非依賴性作用
2、姜黃素在結(jié)直腸癌細(xì)胞中通過ROS激活NRF2
接下來,作者確定姜黃素對CRC細(xì)胞影響的機制。暴露于姜黃素48小時后,HCT116細(xì)胞的ROS水平顯著升高(圖2A)。以ROS誘導(dǎo)劑過氧化叔丁基(tBHP)為陽性對照,姜黃素的作用被抗氧化劑N-乙酰半胱氨酸(NAC)抑制(圖2A,左)。在RKO和SW48細(xì)胞中觀察到類似的結(jié)果。有趣的是,缺乏p53的HCT116細(xì)胞中的ROS水平高于p53存在的HCT116細(xì)胞,這表明p53抑制姜黃素產(chǎn)生ROS(圖2A,右)。值得注意的是,在p53存在和p53缺乏的HCT116細(xì)胞中,姜黃素對細(xì)胞活力的抑制被NAC部分逆轉(zhuǎn)(圖2B)。在RKO和SW48細(xì)胞中觀察到類似的結(jié)果。NAC完全抑制PARP的裂解,從而抑制姜黃素誘導(dǎo)的細(xì)胞凋亡(圖2C)。KEAP1/ NRF2通路是細(xì)胞對氧化應(yīng)激反應(yīng)的中心介質(zhì)。當(dāng)暴露于ROS時,轉(zhuǎn)錄因子NRF2從Keap1釋放,并從細(xì)胞質(zhì)轉(zhuǎn)運到細(xì)胞核,激活其靶基因的轉(zhuǎn)錄。姜黃素在p53存在和p53缺乏的HCT116細(xì)胞中誘導(dǎo)NRF2易位至細(xì)胞核(圖2D)。。暴露于姜黃素后,NRF2蛋白在HCT116細(xì)胞的核部分增加,而在細(xì)胞質(zhì)部分減少(圖2E)。姜黃素不影響NRF2 mRNA的表達(dá)。NQO1是NRF2的保守靶基因,常用于監(jiān)測NRF2通路的活性。與NRF2的激活一致,姜黃素處理后NQO1 mRNA和蛋白的表達(dá)增加(圖2F, G)。重要的是,核易位和NRF2激活的誘導(dǎo)在很大程度上被NAC抑制ROS逆轉(zhuǎn)(圖2D、F、G)。在RKO和SW48細(xì)胞中都能觀察到上述類似的結(jié)果。
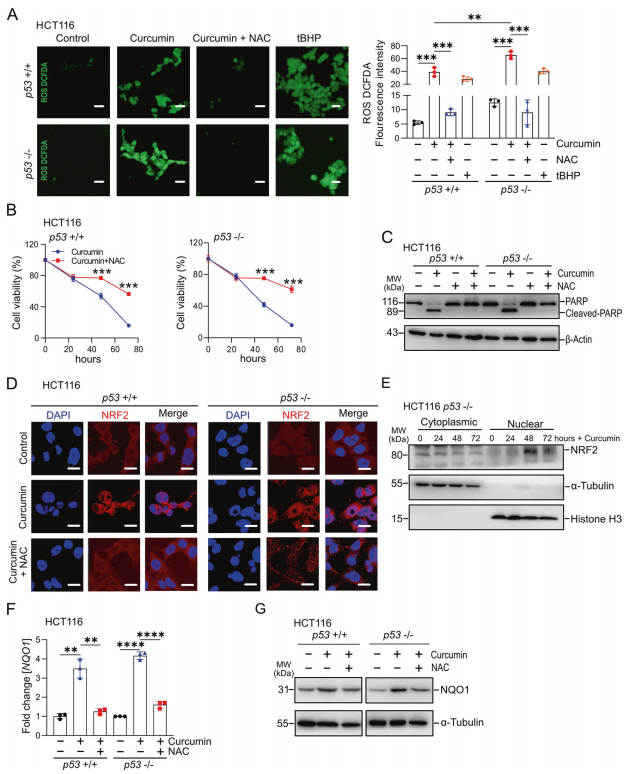
圖2姜黃素在結(jié)直腸癌細(xì)胞中通過ROS激活NRF2
3、姜黃素不依賴于p53來上調(diào)miR-34a和miR-34b/c的表達(dá)
作者先前發(fā)表的觀察結(jié)果表明,miR-34a是由姜黃素誘導(dǎo)的,因此,作者確定了姜黃素在一組p53缺失的等基因CRC細(xì)胞系中誘導(dǎo)miR-34基因是否需要p53。結(jié)果表明,姜黃素誘導(dǎo)了HCT116細(xì)胞中獨立于p53狀態(tài)的pri-miR-34a和pri-miR-34b/c初級轉(zhuǎn)錄本的表達(dá)(圖3A-B)。在CRC細(xì)胞系SW48和RKO中也獲得了類似的結(jié)果(圖3C-F)。在p53存在和p53缺乏的HCT116細(xì)胞中,姜黃素也上調(diào)了成熟miR-34a的水平(圖3G)。由于NAC可以阻止姜黃素對pri-miR34a和pri-miR34b/c的誘導(dǎo),因此作者認(rèn)為它是由ROS介導(dǎo)的(圖3H, I)。綜上所述,這些結(jié)果表明,姜黃素在結(jié)直腸癌細(xì)胞系中以p53獨立和ROS依賴的方式誘導(dǎo)miR-34a和miR-34b/c的表達(dá)。

圖3姜黃素不依賴于p53來上調(diào)miR-34a和miR-34b/c的表達(dá)
4、姜黃素誘導(dǎo)的NRF2直接激活miR-34a和miR-34b/c
接下來,作者假設(shè)被姜黃素誘導(dǎo)的ROS激活的NRF2可能是miR-34a和miR-34b/c表達(dá)的直接誘導(dǎo)因子。事實上,4種不同siRNA對NRF2的抑制阻止了姜黃素在p53缺陷HCT116細(xì)胞中誘導(dǎo)pri-miR-34a和pri-miR-34b/c的表達(dá)。相反,異位NRF2表達(dá)增加了攜帶突變型p53的SW480細(xì)胞中pri-miR-34a和pri-miR-34b/c的表達(dá),以及成熟的miR-34a、NQO-1 mRNA和蛋白的表達(dá)(圖4C- D)。通過檢測miR-34a和miR-34b/c啟動子區(qū)域的基因組序列,作者在miR-34a位點鑒定了三個潛在的NRF2結(jié)合位點(TGAG/CnnnGC),即所謂的ARE(抗氧化反應(yīng)元件),在miR-34b/c位點鑒定了一個潛在的NRF2結(jié)合位點(圖4E-G)。qChIP檢測證實姜黃素處理48小時后,miR-34a和miR-34b/c啟動子內(nèi)四個ARE位點的NRF2占用增強(圖4H)。因此,作者認(rèn)為姜黃素通過激活NRF2誘導(dǎo)miR-34a和miR-34b/c基因的表達(dá),NRF2直接結(jié)合miR-34a和miR-34b/c啟動子并誘導(dǎo)其轉(zhuǎn)錄。并且miR-34的這種激活模式是與p53無關(guān)的。

圖4姜黃素誘導(dǎo)的NRF2直接激活miR-34a和miR-34b/c
5、miR-34a和miR-34b/c介導(dǎo)姜黃素對結(jié)直腸癌細(xì)胞的作用
為了確定姜黃素的作用是否需要誘導(dǎo)miR-34a和miR-34b/c的表達(dá),作者采用了使用CRSPR/CAS9方法使miR-34a和/或miR-34b/c基因缺失的等基因HCT116細(xì)胞。姜黃素處理后,miR34a-或miR-34a/b/c缺陷的HCT116細(xì)胞的活力顯著高于miR-34-存在的HCT116細(xì)胞(圖5A)。與miR-34a/b/c缺失的HCT116細(xì)胞相比,miR-34b/c缺失的細(xì)胞表現(xiàn)出中等的生存能力。與miR-34a/b/c精通的HCT116細(xì)胞相比,miR-34a/b/c缺乏導(dǎo)致姜黃素對細(xì)胞凋亡的增強減弱(圖5B)。在miR-34a/b/c存在的HCT116細(xì)胞中,姜黃素處理誘導(dǎo)了caspase 3的強烈裂解,而在mir -34缺失的細(xì)胞中未檢測到裂解的caspase 3(圖5C)。在miR-34a/b/c存在的HCT116細(xì)胞中,姜黃素抑制了已知的miR-34靶點抗凋亡蛋白Bcl-2的表達(dá),并誘導(dǎo)了促凋亡蛋白BAX的表達(dá)(圖5C)。單獨或聯(lián)合缺失miR-34a和miR-34b/c的HCT116細(xì)胞顯示Bcl-2的表達(dá)升高,姜黃素處理后Bcl-2的表達(dá)超過未處理的WT細(xì)胞,這種影響在miR-34a/b/c缺陷的HCT116細(xì)胞中最為明顯。此外,在miR34a/b/c缺乏的HCT116細(xì)胞中,姜黃素對BAX的誘導(dǎo)作用減弱(圖5C)。因此,miR-34a/b/c對Bcl-2的抑制有助于姜黃素誘導(dǎo)細(xì)胞凋亡。
通過測定在pH 6.0下檢測β-半乳糖苷酶活性來檢測,結(jié)果表明,姜黃素處理后miR-34a、miR-34b/c和miR-34a/b/c缺陷細(xì)胞的衰老沒有增加,而WT HCT116細(xì)胞的衰老明顯增加(圖5D)。最后,姜黃素誘導(dǎo)的對miR-34a、miR-34b/c和miR-34a/b/c缺陷細(xì)胞的遷移和侵襲的抑制明顯低于miR-34存在的HCT116細(xì)胞(圖5E-F)。在所有這些實驗中,聯(lián)合滅活miR-34a和miR-34b/c的細(xì)胞對姜黃素表現(xiàn)出最強的抗性。siRNA敲低NRF2部分阻止了姜黃素抑制細(xì)胞活力(圖5G)、遷移(圖5H)和侵襲(圖5I)。此外,在沒有姜黃素處理的情況下,NRF2的異位表達(dá)抑制了miR-34存在的HCT116細(xì)胞的遷移和侵襲(圖5J-K)。然而,在miR-34a/b/c缺陷的HCT116細(xì)胞中,異位NRF2對遷移和侵襲沒有影響,這表明miR-34a和miR-34b/c是NRF2功能所需的介質(zhì)。綜上所述,作者的研究結(jié)果表明NRF2介導(dǎo)的miR-34a和miR-34b/c基因的激活是姜黃素影響細(xì)胞凋亡、衰老、遷移和侵襲的必要介質(zhì)。
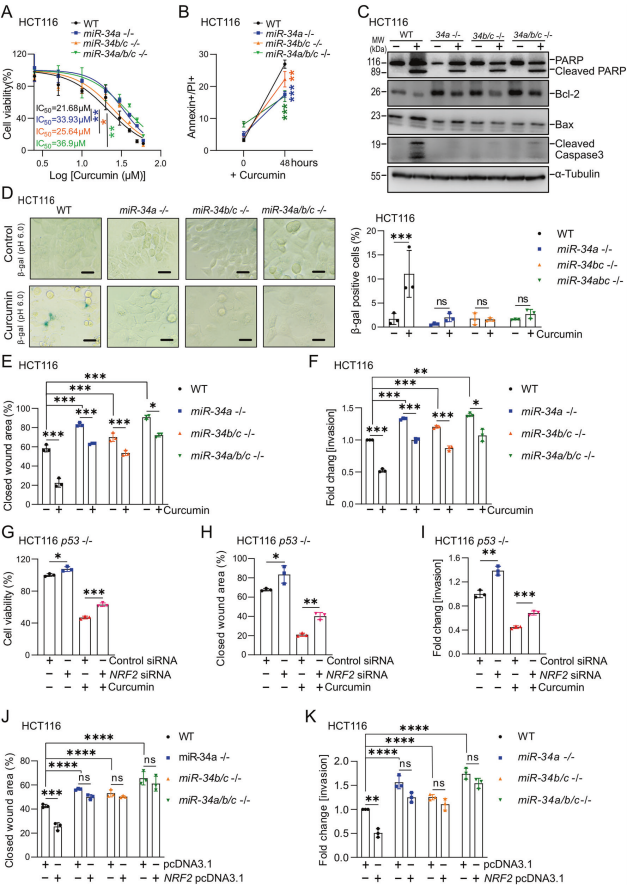
圖5 miR-34a和miR-34b/c介導(dǎo)姜黃素對結(jié)直腸癌細(xì)胞的作用
6、NRF2介導(dǎo)H2O2和tBHP對miR-34a/b/c的誘導(dǎo)
先前有研究表明,H2O2處理誘導(dǎo)的ROS可誘導(dǎo)miR-34a的表達(dá)。因此,作者驗證了這種調(diào)節(jié)是否由NRF2介導(dǎo)。首先,作者分析了H2O2處理后CRC細(xì)胞系的細(xì)胞活力,并測定了表達(dá)p53突變體的HCT116細(xì)胞、p53缺失HCT116細(xì)胞和SW620細(xì)胞的IC50值分別為92.61、48.35和61.52μM。用H2O2或tBHP處理p53存在和p53缺乏的HCT116細(xì)胞以及SW620細(xì)胞,導(dǎo)致NRF2靶基因NQO1的誘導(dǎo),表明H2O2和tBHP激活NRF2(圖6A)。并且,H2O2和tBHP誘導(dǎo)miR-34a/b/c的表達(dá),而siRNA介導(dǎo)的NRF2抑制可以阻止miR-34a/b/c的表達(dá)(圖6B-C)。
7、姜黃素誘導(dǎo)的NRF2逆轉(zhuǎn)缺氧或IL-6對miR-34a/b/c的抑制作用
作者之前已經(jīng)證明缺氧和IL-6介導(dǎo)的STAT3激活抑制了p53缺陷的CRC細(xì)胞中miR-34a/b/c的表達(dá)。為了確定姜黃素是否可以逆轉(zhuǎn)缺氧介導(dǎo)的miR-34a/b/c的抑制,作者在缺氧條件下培養(yǎng)p53缺陷的HCT116細(xì)胞并用姜黃素處理它們。結(jié)果表明,姜黃素完全逆轉(zhuǎn)了缺氧介導(dǎo)的miR-34a/b/c表達(dá)抑制(圖6D-E)。然而,當(dāng)siRNA抑制NRF2時,這種逆轉(zhuǎn)明顯減弱(圖6D-E),表明姜黃素的這種作用在很大程度上是由NRF2激活介導(dǎo)的。另外,IL-6介導(dǎo)的miR-34a和miR-34b/c的抑制被姜黃素完全逆轉(zhuǎn)(圖6F-G)。
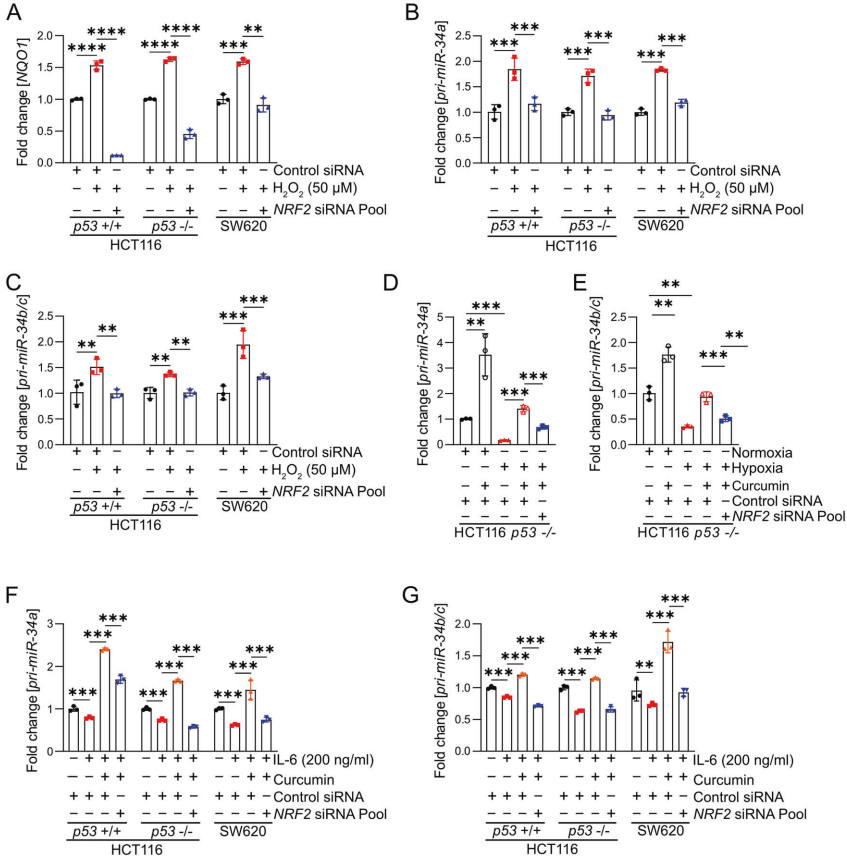
圖6 NRF2軸對H2O2、姜黃素、缺氧和IL-6調(diào)控miR-34a/b/c的影響
8、姜黃素誘導(dǎo)MET并通過誘導(dǎo)miR-34a抑制肺轉(zhuǎn)移形成
接下來,作者打算在移植到小鼠體內(nèi)后研究這些細(xì)胞,作者表征了姜黃素對SW620-Luc2細(xì)胞的影響,SW620-Luc2細(xì)胞穩(wěn)定表達(dá)熒光素酶。結(jié)果表明,姜黃素以劑量依賴性的方式抑制SW620-Luc2細(xì)胞的活力。MTT法測定IC50為14.49 μM。隨后使用了這種濃度的姜黃素,在SW620-Luc2細(xì)胞暴露于姜黃素48小時后,pri-miR-34a和成熟的miR-34a的表達(dá)上調(diào)(圖7A-B)。與成熟的miR34a相比,成熟的miR-34b和miR-34c在暴露于姜黃素的SW620-Luc2細(xì)胞中的表達(dá)水平非常低。因此,作者重點分析了miR-34a在姜黃素對SW620-Luc2細(xì)胞的影響中的作用。作者之前的研究表明,miR-34a在p53誘導(dǎo)的CRC細(xì)胞間充質(zhì)-上皮轉(zhuǎn)化(MET)中起關(guān)鍵作用。因此,作者驗證姜黃素誘導(dǎo)的miR-34a是否可能介導(dǎo)MET。結(jié)果表明,姜黃素處理SW620-Luc2細(xì)胞導(dǎo)致間質(zhì)標(biāo)記物Vimentin (VIM)、SNAIL、SLUG和ZEB1的抑制(圖7E)。此外,姜黃素的這種抑制作用被miR-34a特異性拮抗劑所消除。使用拮抗劑沉默miR-34a來確定姜黃素以miR-34a依賴的方式抑制SW620-Luc2細(xì)胞的入侵和遷移 (圖7C-D)。
最后,作者進(jìn)行了小鼠異種移植實驗,以確定姜黃素是否會影響結(jié)直腸癌細(xì)胞形成肺轉(zhuǎn)移的能力。因此,作者在體外用姜黃素或/和miR-34a特異性拮抗劑處理SW620-Luc2細(xì)胞48小時。隨后,將這些細(xì)胞注射到NOD/SCID小鼠的尾靜脈中,以評估肺轉(zhuǎn)移的形成。縱向無創(chuàng)成像顯示,姜黃素治療SW620-Luc2細(xì)胞在注射后5周內(nèi)完全消除了轉(zhuǎn)移形成(圖7F-G)。然而,拮抗劑同時抑制miR-34a部分恢復(fù)姜黃素治療后的轉(zhuǎn)移形成(圖7F-G)。注射后5周,注射姜黃素處理的SW620-Luc2細(xì)胞的小鼠,切除的肺部沒有宏觀可見的轉(zhuǎn)移(圖7H)。HE染色證實姜黃素處理的細(xì)胞注射小鼠中沒有轉(zhuǎn)移結(jié)節(jié)(圖7H-I)。然而,當(dāng)SW620-luc2細(xì)胞同時接受miR-34anantagomirs和姜黃素處理時,會形成肺轉(zhuǎn)移(圖7H-I)。綜上所述,這些結(jié)果表明姜黃素通過誘導(dǎo)miR-34a抑制轉(zhuǎn)移的形成。

圖7姜黃素誘導(dǎo)MET并通過誘導(dǎo)miR-34a抑制肺轉(zhuǎn)移形成
9、p53和miR-34a/b/c調(diào)節(jié)對姜黃素和/或5-FU的敏感性
最后,作者研究了姜黃素聯(lián)合化療藥物5-氟尿嘧啶(5-FU)對結(jié)直腸癌細(xì)胞活力的影響,該藥物被廣泛用于結(jié)直腸癌的治療。為此,作者使用臨床相關(guān)且可耐受濃度的5-FU (2mg /L)和姜黃素(15μM)。姜黃素和5-FU聯(lián)合處理HCT116細(xì)胞,與單獨處理任何一種化合物相比,顯示出更強的細(xì)胞活力抑制(圖8A)。與存在p53的細(xì)胞相比,缺乏p53的細(xì)胞對姜黃素更敏感,但對5-FU的抗性更強。然而,姜黃素和5-FU處理后,p53缺失細(xì)胞和p53存在細(xì)胞之間沒有差異(圖8A)。與wt細(xì)胞相比,miR-34a-和miR-34a/b/c缺陷細(xì)胞對姜黃素的抗性更強,但對5-FU的抗性僅略強。然而,miR34a-和miR-34a/b/c缺乏導(dǎo)致對姜黃素和5-FU聯(lián)合的抵抗力明顯提高(圖8B)。miR-34a-或miR-34a/b/c缺陷細(xì)胞中p53的進(jìn)一步缺失增加了它們對5-FU的抗性。另外,p53的失活逆轉(zhuǎn)了miR-34a-或miR-34a/b/c缺陷細(xì)胞對姜黃素和姜黃素與5-FU聯(lián)合的耐藥性(圖8C, D)。人結(jié)腸上皮細(xì)胞(HCEC-1CT)和人腸成纖維細(xì)胞(CCD-18Co細(xì)胞)對姜黃素和5-FU的敏感性低于HCT116細(xì)胞(圖8E-F)。綜上所述,這些結(jié)果表明姜黃素可能增強5-FU對CRC細(xì)胞的治療作用。
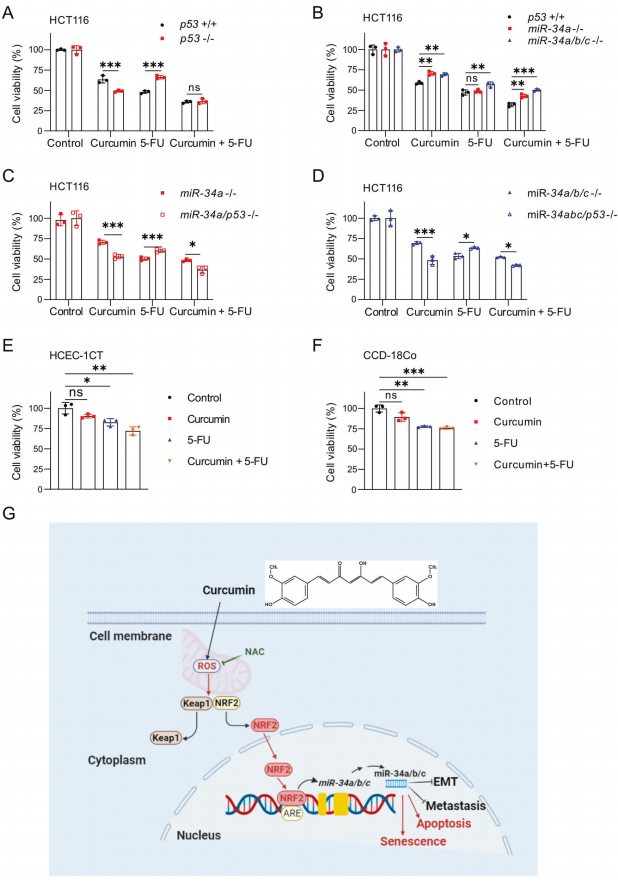
圖8 p53和miR-34a/b/c調(diào)節(jié)對姜黃素和/或5-FU的敏感性
實驗方法
MTT實驗,RNA分離,qRT-PCR,Western blot,染色質(zhì)免疫沉淀反應(yīng),細(xì)胞ROS檢測,異種移植小鼠模型的轉(zhuǎn)移形成,細(xì)胞遷移實驗,細(xì)胞劃痕實驗,RNA敲降,熒光素酶報告實驗
參考文獻(xiàn)
Liu CF, Rokavec M, Huang ZK, Hermeking H. Curcumin activates a ROS/KEAP1/NRF2/miR-34a/b/c cascade to suppress colorectal cancer metastasis. 2023, May. doi: 10.1038/s41418-023-01178-1.