






治療帕金森病——響應(yīng)性人工酶原位持續(xù)供給多巴
口服二羥苯丙氨酸(多巴)補(bǔ)充神經(jīng)元多巴胺仍然是帕金森病(PD)最有效的治療方法。然而,與正常神經(jīng)元持續(xù)穩(wěn)定的多巴胺信號不同,口服多巴引起血漿多巴水平劇烈波動,導(dǎo)致多巴誘導(dǎo)的運(yùn)動障礙。在此,作者報(bào)道了一種基于功能核酸的響應(yīng)性人工酶(FNA-Fe3O4)用于原位連續(xù)生產(chǎn)Dopa。FNA-Fe3O4可以跨越血腦屏障,依靠轉(zhuǎn)鐵蛋白受體適配體靶向病變神經(jīng)元。然后,F(xiàn)NA-Fe3O4響應(yīng)病變神經(jīng)元中過表達(dá)的α -突觸核蛋白mRNA進(jìn)行反義寡核苷酸處理和熒光成像,同時(shí)轉(zhuǎn)換為模擬酪氨酸羥化酶的酪氨酸適配體人工酶(Apt-Fe3O4)用于原位連續(xù)生產(chǎn)多巴。體內(nèi)FNA-Fe3O4治療導(dǎo)致PD小鼠模型中Dopa和多巴胺水平恢復(fù),病理性過表達(dá)的α -突觸核蛋白減少,從而改善運(yùn)動癥狀和記憶缺陷。本文提出的基于功能核酸的響應(yīng)性人工酶策略為PD的診斷和治療提供了一種更神經(jīng)元友好的方法。本研究于2023年5月發(fā)表于期刊《Nature Communications》上,IF:16.6。
技術(shù)路線:
主要研究結(jié)果:
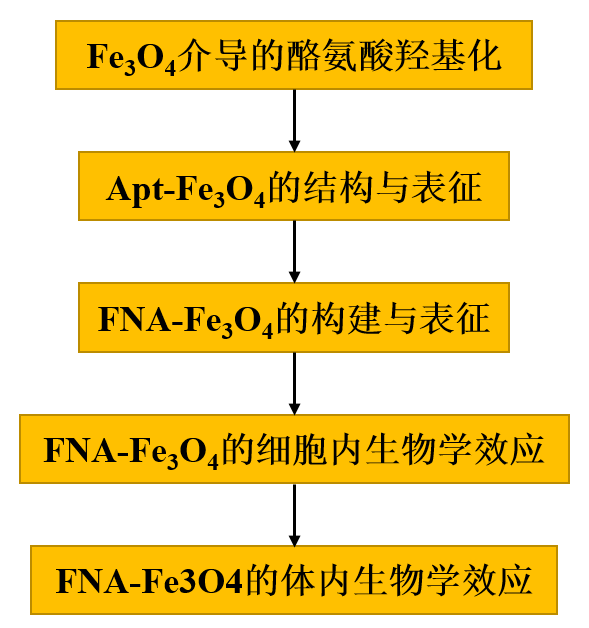
在本研究中,作者首先詳細(xì)解釋了Fe3O4納米粒子在過氧化氫(H2O2)和抗壞血酸(AA)存在的情況下,對酪氨酸羥基化產(chǎn)生多巴(圖1a,Phase I)。進(jìn)一步構(gòu)建了基于酪氨酸適配體的人工酶(Apt-Fe3O4),其中Fe3O4作為催化活性中心,酪氨酸適配體29作為底物結(jié)合位點(diǎn)(圖1a,Phase II)。在病變神經(jīng)元中,F(xiàn)NA-Fe3O4被轉(zhuǎn)化為Apt-Fe3O4以響應(yīng)高水平的SNCA mRNA,從而表現(xiàn)出類似于Dopa產(chǎn)生的酪氨酸羥化酶活性(圖1a,Phase III)。

圖1
1、Fe3O4介導(dǎo)的酪氨酸羥基化
Fe3O4納米顆粒先前被報(bào)道可以催化酪氨酸的多步氧化,通過中間產(chǎn)物多巴(Dopa)生成最終產(chǎn)物多巴色素。然而,中間產(chǎn)品Dopa形成的詳細(xì)機(jī)制尚未得到解釋。因此,作者探究中間產(chǎn)物Dopa的穩(wěn)定存在,以及在病理濃度的AA和H2O2存在下Fe3O4催化酪氨酸羥基化生成Dopa的過程。以市售15 nm水分散表面羧基修飾的Fe3O4納米粒子為模型,考察其催化酪氨酸羥基化合成多巴的性能。圖2a顯示了在10 μg mL-1 Fe3O4、5 mM H2O2和5 mM AA存在下,100 μM酪氨酸羥化轉(zhuǎn)變成多巴過程中的含量變化。反應(yīng)混合物中的每個(gè)組分都是驅(qū)動酪氨酸羥基化為多巴所必需的(圖2b)。根據(jù)這些組分的反應(yīng)活性,可能存在多種活性自由基中間體。tert-Butanol傾向于主要淬滅·OH,而乙醇能有效清除·OH和Fe(IV)=O39。反應(yīng)體系中tert-Butanol的加入在5 min后輕度抑制了反應(yīng)39.4 %,而乙醇的加入誘導(dǎo)了更嚴(yán)重的抑制60.4 %(圖2c)。這些差異可歸因于羥基化過程中·OH和高自旋Fe(IV)=O的參與。
為進(jìn)一步了解Fe3O4在H2O2和AA存在下催化酪氨酸羥基化生成Dopa的機(jī)理,作者進(jìn)行了一系列電子自旋共振(ESR)實(shí)驗(yàn),并將這些結(jié)果與反應(yīng)的活性中間體進(jìn)行了關(guān)聯(lián)。圖2d顯示了Fe3O4與H2O2共存的混合體系中明顯的羥基自由基(·OH)信號,這與Fe3O4納米酶40固有的過氧化物酶活性一致。圖2e表明,F(xiàn)e3O4與AA共存時(shí),沒有檢測到自由基的產(chǎn)生。而在Fe3O4、H2O2和AA存在下,觀察到新的自由基信號為過氧自由基(·OOH),并且伴隨著體系中·OH的完全消失(圖2f)。H2O2與AA之間沒有發(fā)生直接的自由基反應(yīng)(圖2g)。這些結(jié)果表明AA是·OOH形成的關(guān)鍵促進(jìn)劑并導(dǎo)致·OH耗竭。進(jìn)一步地,在·OOH的混合體系中加入酪氨酸底物后,·OOH的強(qiáng)度降低,自由基種類沒有發(fā)生變化(圖2h),暗示·OOH為酪氨酸向多巴的轉(zhuǎn)化提供了關(guān)鍵物種。據(jù)報(bào)道,·OOH與Fe2+結(jié)合生成高自旋Fe(IV)= O中間體41,其結(jié)構(gòu)與生物蛋白酪氨酸羥化酶的活性中心相似。以二甲基亞砜(DMSO)作為Fe(IV)=O的探針,通過氧原子轉(zhuǎn)移與Fe(IV)= O反應(yīng)生成二甲基砜(MSM)。根據(jù)圖2i的氣相色譜-質(zhì)譜(GC-MS)分析結(jié)果,隨著反應(yīng)時(shí)間的延長,MSM的峰逐漸增強(qiáng),表明可能發(fā)生了Fe(IV)= O。這些結(jié)果為酪氨酸羥基化的機(jī)理路徑提供了有效的解釋(圖2j)。Fe3O4與H2O2反應(yīng)產(chǎn)生的·OH在AA的催化作用下轉(zhuǎn)化為·OOH。然后·OOH介導(dǎo)生成活性中間體Fe(IV)=O,最后羥基化酪氨酸生成多巴。

圖2 Fe3O4納米顆粒的酪氨酸羥化酶模擬活性
2、Apt-Fe3O4的結(jié)構(gòu)與表征
適體結(jié)合位點(diǎn)與納米酶的共價(jià)連接被引入作為一種通用的方法,通過將反應(yīng)底物集中在催化納米酶核心上來提高納米酶的催化活性,從而模擬天然酶的結(jié)合和催化活性位點(diǎn)功能(圖3a)。在確定Fe3O4納米酶的酪氨酸羥化酶模擬活性后,通過EDC/NHS介導(dǎo)的酰胺縮合反應(yīng)將Fe3O4納米酶與5 '端氨基修飾的酪氨酸適體共價(jià)偶聯(lián),構(gòu)建基于酪氨酸適體的人工酶Apt-Fe3O4。作者還制備了隨機(jī)序列DNA偶聯(lián)的Fe3O4(Ran-Fe3O4)用于對比。
作者考察不同濃度的酪氨酸(所有實(shí)驗(yàn)均使用5 mM H2O2和5 mM AA)在Fe3O4、Ran-Fe3O4和Apt-Fe3O4(10 μg mL-1)的催化作用下,酪氨酸羥化生成多巴的速率隨酪氨酸濃度的變化,如圖3b的動力學(xué)曲線所示。對于Apt-Fe3O4或?qū)φ眨^察到Michaelis-Menten-type saturation kinetic曲線。與純Fe3O4相比,Ran-Fe3O4的催化活性幾乎相同,而Apt-Fe3O4的催化活性顯著提高。Apt-Fe3O4催化活性的增強(qiáng)歸因于靠近催化界面的底物的高局部濃度,這是由于適配體與酪氨酸底物的特異性結(jié)合。
值得注意的是,水溶液作為最簡單的模型,無法模擬實(shí)際的細(xì)胞內(nèi)大分子擁擠環(huán)境。因此,在以20 wt % PEG-20000作為擁擠劑的模擬擁擠環(huán)境中評估了Apt-Fe3O4和對照的催化能力。如圖3c所示,與水溶液中的催化速率相比,F(xiàn)e3O4和Ran-Fe3O4的催化速率明顯下降,表明它們的催化功能在復(fù)雜的細(xì)胞環(huán)境中受到嚴(yán)重干擾。然而,Apt-Fe3O4令人滿意地保留了其大部分催化效率,這無疑是適配體對酪氨酸的選擇性捕獲。酪氨酸與適配體的特異性結(jié)合,增加了酪氨酸在Apt-Fe3O4上的停留時(shí)間,有利于活性中間體的形成,使反應(yīng)易于在擁擠的環(huán)境中進(jìn)行。這一特性有利于基于酪氨酸適配體的人工酶Apt-Fe3O4在復(fù)雜的細(xì)胞內(nèi)環(huán)境中執(zhí)行催化任務(wù)。
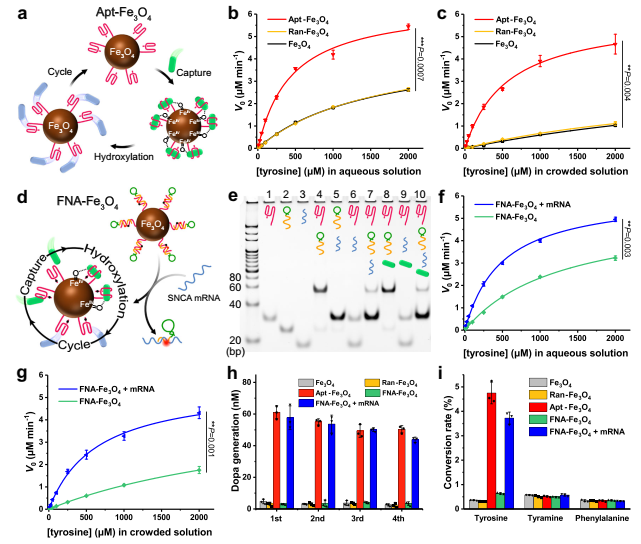
圖3 人工酶的表征及其催化和熒光成像性能
3、FNA-Fe3O4的構(gòu)建與表征
其次,為實(shí)現(xiàn)人工酶對PD治療的高效BBB穿越和神經(jīng)元特異性響應(yīng),設(shè)計(jì)了由SNCA反義寡核苷酸和TfR適配體組成的阻斷鏈,并與Apt-Fe3O4上的酪氨酸適配體雜交,構(gòu)建了基于功能核酸的響應(yīng)型人工酶FNA-Fe3O4(圖3D)。
通過聚丙烯酰胺凝膠電泳(PAGE)驗(yàn)證FNA-Fe3O4響應(yīng)SNCA mRNA的鏈置換反應(yīng)性。如圖3e所示,嵌段鏈可以首先與酪氨酸適配體雜交(泳道4)。該雙鏈在酪氨酸的生理濃度范圍內(nèi)穩(wěn)定(泳道8)。然而,嵌段鏈隨后被SNCA mRNA取代以釋放游離的酪氨酸適體(泳道10)。同時(shí),監(jiān)測SNCA相關(guān)生物標(biāo)志物的含量可以為PD的熒光成像提供有效手段。
隨后測定了FNA-Fe3O4在SNCA mRNA存在和不存在時(shí)的酪氨酸羥化酶模擬活性。如圖3f中的動力學(xué)曲線所示,在水溶液和擁擠溶液中,F(xiàn)NA-Fe3O4與SNCA mRNA發(fā)生鏈置換反應(yīng)后表現(xiàn)出較高的催化活性。而單獨(dú)的FNA-Fe3O4的催化活性保持在較低的水平,尤其是在擁擠溶液中。這一特性提供FNA-Fe3O4不會影響正常細(xì)胞中多巴胺穩(wěn)態(tài)的潛力,僅在SNCA mRNA異常過表達(dá)的細(xì)胞中提供Dopa。
FNA-Fe3O4與SNCA mRNA反應(yīng)后,能夠連續(xù)捕獲酪氨酸溶液中的酪氨酸,并在含有H2O2和AA的溶液中催化酪氨酸羥基化,反應(yīng)5 min內(nèi)的多巴生成在4次重復(fù)中保持穩(wěn)定(圖3h),這將是人工酶典型的循環(huán)催化過程的有力證明。此外,以幾種常見的能夠芳香環(huán)羥基化的胞內(nèi)小分子(酪胺和苯丙氨酸)作為模型干擾物,測試人工酶對酪氨酸的底物選擇性。如圖3i所示,在反應(yīng)5 min內(nèi),F(xiàn)NA-Fe3O4 + mRNA對酪氨酸的羥基化轉(zhuǎn)化率遠(yuǎn)高于其他干擾物質(zhì),對酪氨酸表現(xiàn)出優(yōu)異的選擇性,證明了其胞內(nèi)抗干擾能力。此外,在含有酪氨酸、H2O2和AA的Apt-Fe3O4、FNA-Fe3O4和FNA-Fe3O4 + mRNA(10 μg mL-1)溶液中,反應(yīng)30 min后均未產(chǎn)生DNA片段,說明了反應(yīng)過程中DNA鏈的穩(wěn)定性。
4、FNA-Fe3O4的細(xì)胞內(nèi)生物學(xué)效應(yīng)
BBB是FNA-Fe3O4進(jìn)入多巴胺能神經(jīng)元的第一個(gè)障礙。因此,用表達(dá)TfR的永生化小鼠腦內(nèi)皮細(xì)胞(bEnd.3)建立體外BBB模型,在頂端側(cè)加入人工酶,考察其穿膜效率(圖4a)。隨著時(shí)間的推移,F(xiàn)NA-Fe3O4在基底側(cè)逐漸累積,120 min后轉(zhuǎn)運(yùn)效率上升至21.8 %(圖4b)。相比之下,Apt-Fe3O4,Ran-Fe3O4,和不含TfR適配體的Fe3O4在基底側(cè)緩慢積累。這些結(jié)果表明FNA-Fe3O4可以高效地通過體外BBB模型,其中TfR適配體發(fā)揮了重要作用。
在驗(yàn)證了FNA-Fe3O4的跨BBB作用后,作者進(jìn)一步檢測其在神經(jīng)元中的內(nèi)化情況。作者使用TfR陽性的小鼠小膠質(zhì)細(xì)胞BV2細(xì)胞作為PD的細(xì)胞模型。以Cy5修飾的不含BHQ2的FNA-Fe3O4為熒光探針,以Apt-Fe3O4和Ran-Fe3O4為熒光探針。與不含TfR適配體的Apt-Fe3O4和Ran-Fe3O4相比,F(xiàn)NA-Fe3O4對目標(biāo)BV2細(xì)胞具有更好的結(jié)合位移(圖4c)。代表FNA-Fe3O4的強(qiáng)烈Cy5熒光信號主要定位于BV2細(xì)胞的細(xì)胞質(zhì)中,并在較長時(shí)間內(nèi)保持在胞內(nèi)高水平(圖4d)。細(xì)胞活性測試結(jié)果表明FNA-Fe3O4對BV2細(xì)胞幾乎沒有明顯的細(xì)胞毒性(圖4e),表明其與腦內(nèi)皮細(xì)胞具有良好的生物相容性。這些性質(zhì)為FNA-Fe3O4的細(xì)胞內(nèi)應(yīng)用提供基礎(chǔ)。
為評估核酸組分在FNA-Fe3O4上的功能,作者設(shè)計(jì)了兩種對照納米材料。Cat-Fe3O4代表酪氨酸羥化酶模擬活性的影響,ASO-Fe3O4代表反義寡核苷酸療法(圖5a-b)。作者通過其脫羧產(chǎn)物多巴胺的含量來監(jiān)測。如圖5c所示,F(xiàn)NA-Fe3O4處理的正常細(xì)胞內(nèi)多巴胺含量變化不大,表明其不會干擾正常神經(jīng)元內(nèi)多巴胺穩(wěn)態(tài)。在FNA-Fe3O4處理的PD細(xì)胞中,多巴胺含量在8 h內(nèi)呈逐漸上升趨勢,隨后達(dá)到穩(wěn)定平臺期。這一結(jié)果證明,F(xiàn)NA-Fe3O4可以在細(xì)胞中發(fā)揮類似酪氨酸羥化酶的功能,以應(yīng)對細(xì)胞特異性空間控制的SNCA mRNA的異常過度表達(dá)。同時(shí),Cat-Fe3O4也表現(xiàn)出類似的酪氨酸羥化酶模擬活性,而ASO-Fe3O4則沒有,這反映了酪氨酸適配體對人工酶催化的決定性作用。作者還嘗試了直接多巴孵育(0.5、2、8 μM)對細(xì)胞內(nèi)多巴胺水平的影響。孵育2 h后更換培養(yǎng)液模擬多巴胺短的血漿半衰期,細(xì)胞內(nèi)多巴胺含量逐漸降低(圖5d)。較高劑量的多巴維持了較長時(shí)間的多巴胺水平,但也導(dǎo)致了多巴胺含量更劇烈的波動。總的來說,多巴胺水平的長期增加意味著多巴治療的時(shí)間更長,而多巴胺水平的劇烈波動預(yù)示著神經(jīng)元中多巴胺的穩(wěn)態(tài)失調(diào)。這是導(dǎo)致多巴的治療效果和副作用控制無法平衡的根本原因。相比之下,F(xiàn)NA-Fe3O4提供的原位連續(xù)多巴產(chǎn)量表現(xiàn)出更長的持續(xù)時(shí)間和更可接受的多巴胺波動。需要指出的是,由于多巴和多巴胺的有效生理濃度遠(yuǎn)低于酪氨酸,因此應(yīng)用FNA-Fe3O4進(jìn)行酪氨酸羥基化不會影響細(xì)胞內(nèi)酪氨酸穩(wěn)態(tài)。
將阻斷鏈與SNCA mRNA雜交也降低了SNCA的表達(dá)。人工酶共孵育36 h后,F(xiàn)NAFe3O4和ASO-Fe3O4降低PD細(xì)胞模型SNCA蛋白表達(dá)水平(圖5e),有利于延緩神經(jīng)元退變。同在圖5f的熒光分析中,顯示EGFP綠色熒光信號的細(xì)胞為SNCA mRNA過表達(dá)細(xì)胞。FNA-Fe3O4對共培養(yǎng)的正常細(xì)胞和PD細(xì)胞顯示出不同的熒光信號強(qiáng)度,證明FNA-Fe3O4可以響應(yīng)過表達(dá)的SNCA mRNA通過熒光成像區(qū)分正常和PD神經(jīng)元(圖5g)。相應(yīng)的流式細(xì)胞儀獲得了一致的熒光分析結(jié)果(圖5h)。這些結(jié)果表明FNA-Fe3O4能夠監(jiān)測細(xì)胞內(nèi)SNCA mRNA的表達(dá)水平。

圖4 人工酶的細(xì)胞可利用性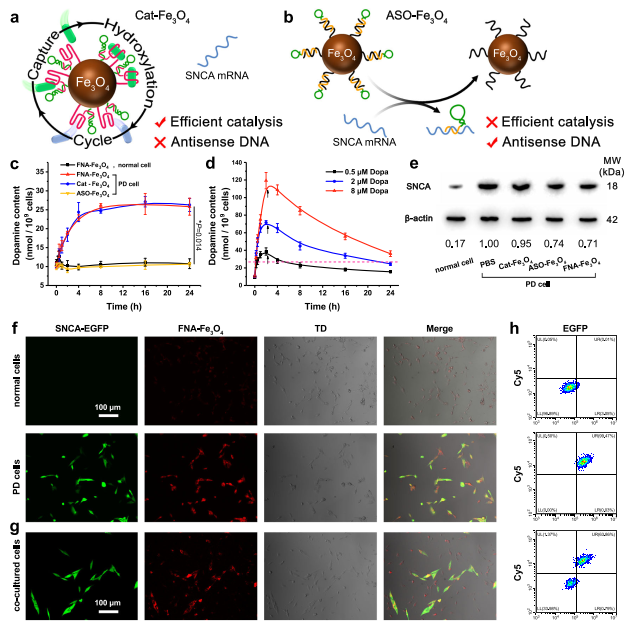
圖5 人工酶的細(xì)胞內(nèi)生物學(xué)效應(yīng)
5、FNA-Fe3O4的體內(nèi)生物學(xué)效應(yīng)
研究分別以野生型小鼠和攜帶小鼠胸腺細(xì)胞抗原1(Thy1)啟動子驅(qū)動的人野生型SNCA的Thy1-SNCA轉(zhuǎn)基因小鼠作為正常和PD動物模型。與正常小鼠相比,尾靜脈注射FNA-Fe3O4后4 h內(nèi),PD小鼠腦內(nèi)可觀察到逐漸增強(qiáng)的Cy5熒光信號(圖6a),表明FNA-Fe3O4可實(shí)現(xiàn)PD動物模型的成像。
隨后,在PD小鼠尾靜脈注射Cat-Fe3O4、ASO-Fe3O4或FNA-Fe3O4(4 mg kg-1)時(shí)進(jìn)行紋狀體微透析,分析人工酶催化原位Dopa產(chǎn)生。具有類酪氨酸羥化酶特性的Cat-Fe3O4和FNA-Fe3O4使腦組織中多巴和多巴胺水平升高至略低于正常水平,而ASO-Fe3O4未引起多巴和多巴胺水平的變化(圖6b-c)。這一趨勢表明人工酶介導(dǎo)了腦內(nèi)多巴胺系列神經(jīng)遞質(zhì)生成的恢復(fù)。相應(yīng)地,比較了Dopa給藥期間的微透析結(jié)果。多巴胺的上升趨勢只維持了(約4 h內(nèi))短暫的時(shí)間且波動較大(圖6d),從而顯示出人工酶策略的優(yōu)越性。經(jīng)過20天的治療后(圖6e),反義寡核苷酸ASO-Fe3O4和FNA-Fe3O4顯著降低了腦組織中SNCA的表達(dá),而Cat-Fe3O4對過表達(dá)SNCA的影響可以忽略(圖6f)。這一結(jié)果揭示人工酶上反義寡核苷酸部分對于調(diào)控過表達(dá)SNCA的重要性。
持續(xù)的原位多巴生成和緩解異常的SNCA過表達(dá)共同作用于Thy1-SNCA小鼠的PD病理特征。因此,進(jìn)一步通過爬桿實(shí)驗(yàn)和懸線實(shí)驗(yàn)(圖6g-h)評價(jià)小鼠的運(yùn)動協(xié)調(diào)能力。可見,PD小鼠的運(yùn)動協(xié)調(diào)能力明顯差于健康小鼠。Cat-Fe3O4和ASO-Fe3O4部分恢復(fù)了PD小鼠的運(yùn)動能力,分別反映了酪氨酸羥化酶替代治療和反義寡核苷酸治療對PD癥狀的緩解。單獨(dú)使用Cat-Fe3O4的治療效果略優(yōu)于ASO-Fe3O4,因?yàn)樵欢喟凸┙o可以直接作用于多巴胺能神經(jīng)系統(tǒng)。反義寡核苷酸治療的貢獻(xiàn)在于緩解PD病理過程,因此FNA-Fe3O4治療的小鼠表現(xiàn)出比單藥治療更好的性能。此外,PD小鼠可表現(xiàn)出學(xué)習(xí)記憶能力的下降,可通過被動回避實(shí)驗(yàn)進(jìn)行檢測。24 h滯留中更長的穿越潛伏期代表更完整的學(xué)習(xí)記憶。在PD小鼠中給予Cat-Fe3O4和ASO-Fe3O4在相似的水平上有助于改善學(xué)習(xí)和記憶的下降,并且FNA-Fe3O4治療更明顯。這些結(jié)果表明FNA-Fe3O4可以促進(jìn)PD小鼠模型的運(yùn)動障礙和記憶缺陷的恢復(fù),其治療作用來源于酪氨酸羥化酶模擬活性和反義寡核苷酸的協(xié)同作用。
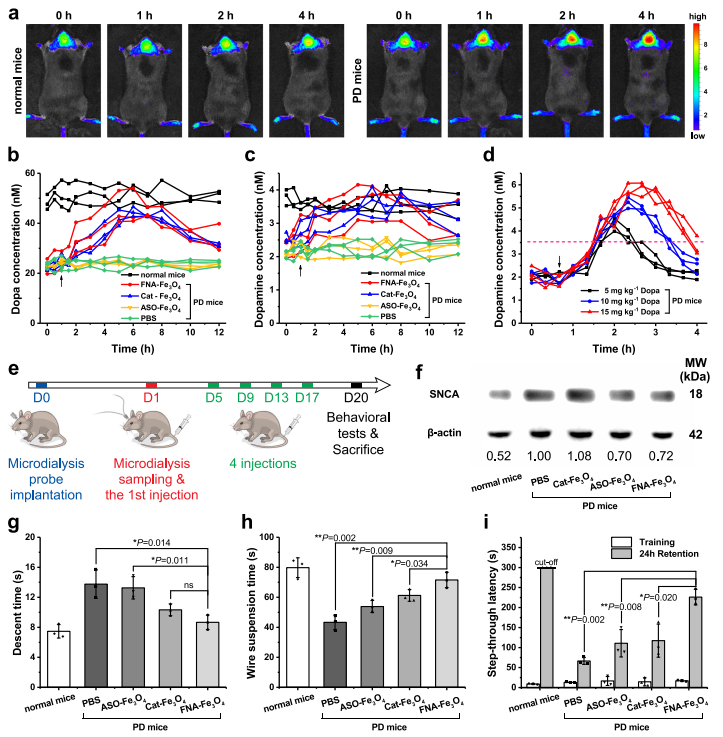
圖6 人工酶的體內(nèi)生物學(xué)效應(yīng)
實(shí)驗(yàn)方法
SH-SY5Y和bEnd.3細(xì)胞培養(yǎng),體外構(gòu)建血腦屏障細(xì)胞模型,人工酶的細(xì)胞內(nèi)化,CCK-8,細(xì)胞內(nèi)多巴胺和酪氨酸含量的測定,qRT-PCR,免疫印跡,人工酶對細(xì)胞SNCA mRNA的熒光分析,C57BL/6 N(6個(gè)月)小鼠,Thy1-SNCA 轉(zhuǎn)基因小鼠,F(xiàn)NA-Fe3O4中塊鏈的質(zhì)譜,離體成像,體內(nèi)血液循環(huán)測定,免疫熒光,血清生化分析,溶血試驗(yàn)
參考文獻(xiàn)
Fang X, Yuan M, Zhao F, Yu A, Lin Q, Li S, Li H, Wang X, Yu Y, Wang X, Lin Q, Lu C, Yang H. In situ continuous Dopa supply by responsive artificial enzyme for the treatment of Parkinson's disease. Nat Commun. 2023 May 9;14(1):2661. doi: 10.1038/s41467-023-38323-w. PMID: 37160866; PMCID: PMC10169781.