






單細(xì)胞轉(zhuǎn)錄組學(xué)闡明了嗜堿性粒細(xì)胞分化軌跡, 并鑒定了成熟嗜堿性粒細(xì)胞上游的前嗜堿性粒細(xì)胞
嗜堿性粒細(xì)胞是最稀有的粒細(xì)胞,被認(rèn)為是2型免疫反應(yīng)的關(guān)鍵細(xì)胞。然而,它們的分化路徑仍未完全闡明。在這項(xiàng)研究中,我們通過單細(xì)胞RNA測序分析來評估嗜堿性粒細(xì)胞的個體發(fā)育軌跡。結(jié)合流式細(xì)胞術(shù)和功能分析,我們發(fā)現(xiàn)c-Kit-CLEC12Ahi前嗜堿性粒細(xì)胞位于前嗜堿性粒細(xì)胞和肥大細(xì)胞祖細(xì)胞(pre-BMPs)下游和CLEC12Alo成熟嗜堿性粒細(xì)胞上游。轉(zhuǎn)錄分析預(yù)測,在基因表達(dá)譜方面,前嗜堿性粒細(xì)胞群體包括先前定義的嗜堿性粒細(xì)胞祖(BaP)-like 細(xì)胞。與成熟嗜堿性粒細(xì)胞相比,前嗜堿性粒細(xì)胞具有較高的增殖能力,對非IgE刺激的反應(yīng)較好,但對抗原加IgE刺激的反應(yīng)較弱。盡管前嗜堿性粒細(xì)胞通常停留在骨髓中,但它們會在蛔蟲感染的組織中出現(xiàn),可能是通過IL-3介導(dǎo)抑制其在骨髓中的滯留。因此,本研究確定了在嗜堿性粒細(xì)胞個體發(fā)育過程中連接pre-BMPs和成熟嗜堿性粒細(xì)胞之間的前嗜堿性粒細(xì)胞。
該研究于2023年5月發(fā)表發(fā)表在《Nature communications》,IF:17.694。
技術(shù)路線:

實(shí)驗(yàn)方法:抗體、細(xì)胞培養(yǎng)、流式細(xì)胞儀分析與細(xì)胞分選、Giemsa染色、scRNA-seq、bulk RNA-seq分析、蠕蟲感染、嗜堿性粒細(xì)胞分類純化后的體外細(xì)胞培養(yǎng)、統(tǒng)計(jì)分析。
1、骨髓來源的嗜堿性粒細(xì)胞(BMBAs)包含兩個亞群
嗜堿性粒細(xì)胞是數(shù)量最少、壽命較短的粒細(xì)胞,因此常常使用骨髓來源的嗜堿性粒細(xì)胞(BMBAs)作為新鮮動物嗜堿性粒細(xì)胞的替代物。為了確定BMBAs的最佳培養(yǎng)條件,我們將小鼠骨髓細(xì)胞與不同濃度的IL-3進(jìn)行培養(yǎng),濃度范圍從0到100 ng/mL,培養(yǎng)時間為7天。隨著IL-3濃度增加至0.3 ng/mL,培養(yǎng)物中CD200R3+ c-Kit?嗜堿性粒細(xì)胞的比例增加到約60%。在較高濃度的IL-3下,嗜堿性粒細(xì)胞的比例反而下降,而CD200R3+ c-Kit+肥大細(xì)胞的比例增加。因此,0.3 ng/mL的IL-3被證明是制備高純度嗜堿性細(xì)胞的最佳選擇,雖然遠(yuǎn)低于通常使用的濃度。
在上述實(shí)驗(yàn)中,我們注意到CD200R3+ c-Kit? BMBAs中存在兩個亞群:FcεRIαhiCD49blo和FcεRIαloCD49bhi(圖1a)。不論使用C57BL/6還是BALB/c小鼠,前者亞群的頻率隨著IL-3濃度增加而下降,表明IL-3對這兩個亞群之間的平衡產(chǎn)生影響。值得注意的是,F(xiàn)cεRIαloCD49bhi BMBAs顯示出與外周血嗜堿性粒細(xì)胞相同的環(huán)狀細(xì)胞核,而FcεRIαhiCD49blo BMBAs則顯示出非典型形態(tài),呈腎形凹陷狀,胞體較大(圖1b)。FcεRIαhiCD49blo BMBas由于細(xì)胞體較大,其前向散射強(qiáng)度高于FcεRIαloCD49bhi BMBas(圖1c)。
為了進(jìn)一步表征和比較這兩個BMBAs亞群,我們對骨髓細(xì)胞的IL-3培養(yǎng)物中的細(xì)胞進(jìn)行了無偏向單細(xì)胞RNA測序(scRNA-seq)分析。通過Seurat聚類分析scRNA-seq數(shù)據(jù)集,確定了11個細(xì)胞簇,其中簇0、簇2和簇10是根據(jù)Cd200r3和Mcpt8的高表達(dá)以及Kit的低表達(dá)被判定為嗜堿性粒細(xì)胞。在這三個簇中(分別在圖1d中表示為BMBA1、BMBA2和BMBA3),BMBA2和BMBA3的Fcer1a(編碼FcεRIα)表達(dá)較低,而Itga2(編碼CD49b)表達(dá)較高,相比之下BMBA1則相反(圖1d、e)。因此,BMBA2和BMBA3似乎對應(yīng)于FcεRIαloCD49bhi BMBAs,而BMBA1對應(yīng)于FcεRIαhiCD49blo BMBAs。在BMBA1和BMBA2/3之間,我們檢測到5382個差異表達(dá)基因(DEGs),其中4811個基因在BMBA1中上調(diào),而571個基因在BMBA2/3中上調(diào)(圖1f)。其中,我們選擇了兩個編碼細(xì)胞表面蛋白的基因,Clec12a編碼CLEC12A和Cd9編碼CD9(圖1e、f),作為可能的代用標(biāo)記物,用于在后續(xù)實(shí)驗(yàn)中區(qū)分BMBAs的兩個亞群。使用特異性單克隆抗體進(jìn)行的流式細(xì)胞術(shù)分析顯示,F(xiàn)cεRIαhiCD49blo和FcεRIαloCD49bhi BMBAs幾乎對應(yīng)于CLEC12AhiCD9lo和CLEC12AloCD9hi BMBAs(圖1g)。同樣,CLEC12AhiCD9lo和CLEC12AloCD9hi BMBAs幾乎對應(yīng)于FcεRIαhiCD49blo和FcεRIαloCD49bhi BMBAs(圖1h)。
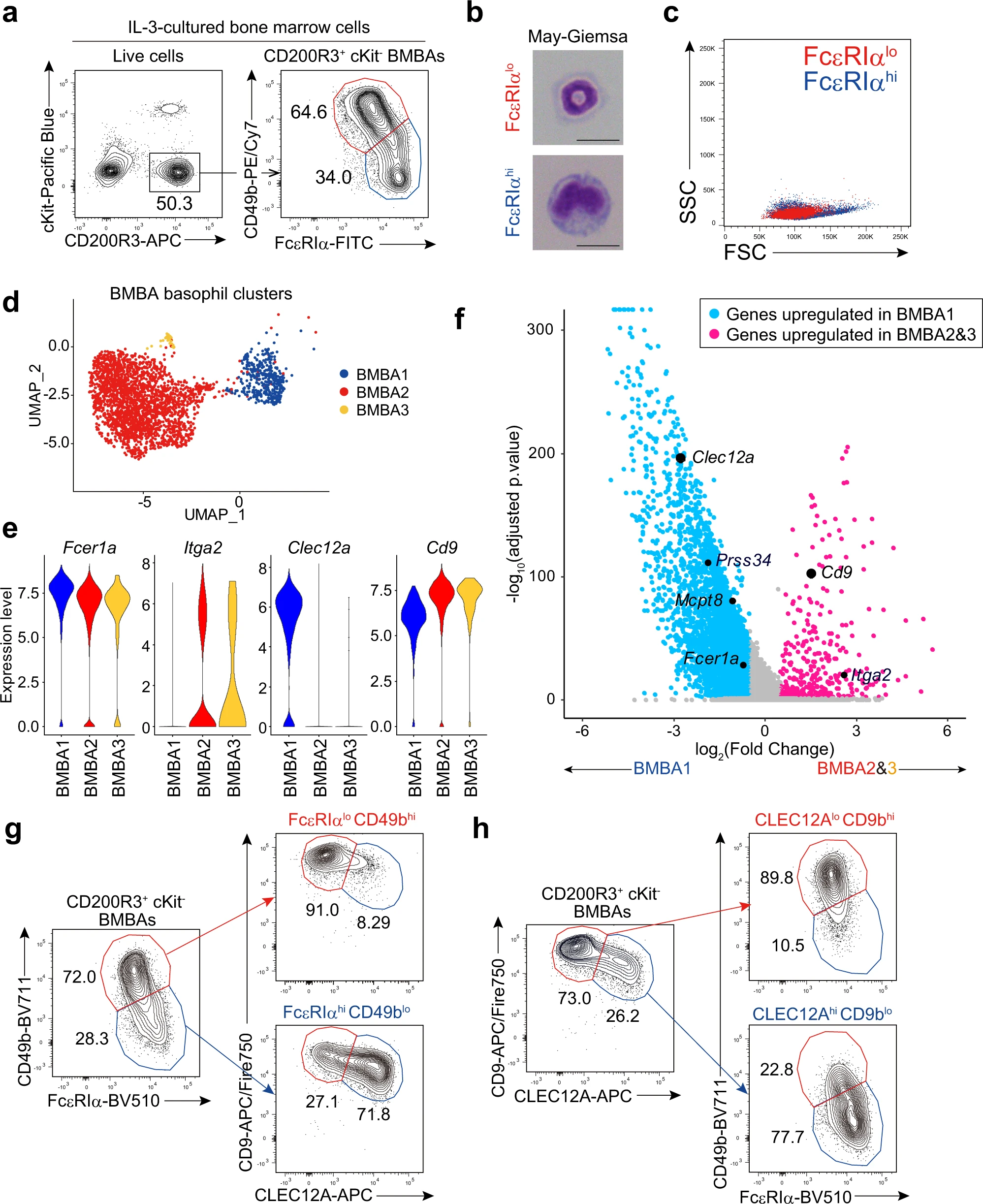
圖1|骨髓源性嗜堿性粒細(xì)胞(BMBA)由兩個不同的亞群組成。
2、在骨髓中檢測到兩個嗜堿性粒細(xì)胞的亞群,而在外周血和脾臟中只檢測到其中的一個亞群
接下來,我們檢查了是否可以在從小鼠中新鮮分離的嗜堿性粒細(xì)胞中檢測到在BMBAs中鑒定出的兩個嗜堿性粒細(xì)胞亞群。在c-Kit-CD200R3+ CD49b+嗜堿性粒細(xì)胞中,F(xiàn)cεRIαloCD49bhi/CLEC12AloCD9hi亞群在外周血、脾臟和骨髓中都很容易被鑒定出來(圖2a)。相反,盡管骨髓中存在,但在外周血和脾臟中很少檢測到FcεRIαhiCD49blo /CLEC12AhiCD9lo亞群(圖2a)。骨髓、脾臟和外周血中的CLEC12AloCD9hi嗜堿性粒細(xì)胞具有典型的環(huán)形或雙裂核,而骨髓中的CLEC12AhiCD9lo嗜堿性粒細(xì)胞則顯示出具有腎臟形狀的凹陷核和較大的細(xì)胞體,并在流式細(xì)胞術(shù)中顯示出較高的前向散射(FSC),這與BMBAs中觀察到的情況相符(圖2b、c)。值得注意的是,Lin-cKit-CD34+ CD200R3+ BaPs也顯示出CLEC12AhiCD9lo表型,而在cKit-CD200R3+嗜堿性粒細(xì)胞譜系細(xì)胞中,約10%的CLEC12Ahi亞群CD34表達(dá)量較低(圖2d),這表明CLEC12Ahi嗜堿性粒細(xì)胞可能包含單能的類似BaP的群體。支持這一觀點(diǎn)的是,CD34+ BaPs和CD34- CLEC12Ahi嗜堿性粒細(xì)胞顯示出類似的形態(tài)和表面表達(dá)特征也支持了這一觀點(diǎn)。

圖2|在骨髓中也檢測到兩個亞群的嗜堿性粒細(xì)胞,而在外周血和脾中只檢測到一個亞群
3、單細(xì)胞RNA測序(scRNA-seq)分析鑒定出骨髓嗜堿性粒細(xì)胞中的4個簇
我們使用Mcpt8GFP轉(zhuǎn)基因小鼠進(jìn)行分析,該轉(zhuǎn)基因小鼠在Mcpt8啟動子/增強(qiáng)子下表達(dá)GFP編碼基因,以有效富集和分離稀有的嗜堿性粒細(xì)胞,以分析骨髓嗜堿性粒細(xì)胞的分化軌跡。我們確認(rèn),CLEC12Ahi和CLEC12Alo嗜堿性粒細(xì)胞亞群中的大多數(shù)細(xì)胞都表達(dá)GFP。從骨髓和脾臟中分離的Lineage(Lin)- GFP+細(xì)胞分別進(jìn)行了單細(xì)胞RNA測序分析。單細(xì)胞RNA測序數(shù)據(jù)的聚類分析鑒定出12個簇,其中0、1和4號簇對應(yīng)于KitCd200r3+ Mcpt8+嗜堿性粒細(xì)胞。對這三個簇進(jìn)行重新聚類,鑒定出1個Fcer1a+ Kit+Cd34+類似于pre-BMPs細(xì)胞的簇,1個Clec12ahi嗜堿性粒細(xì)胞簇(Baso1)和2個Clec12alo嗜堿性粒細(xì)胞簇(Baso2和Baso3)(圖2e、f)。與流式細(xì)胞術(shù)分析結(jié)果一致,Clec12ahi嗜堿性粒細(xì)胞(Baso1)在骨髓中豐富,而在脾臟中較少(圖2e、f)。與流式細(xì)胞術(shù)分析一致,21%的Clec12ahi Baso1細(xì)胞的Cd34表達(dá)水平較低(圖2f)。
為了驗(yàn)證我們的發(fā)現(xiàn),我們重新分析了由Weinreb等人報(bào)道的公開可用的scRNA-seq數(shù)據(jù)集(GEO登錄號:GSE140802),鑒定出1個Fcer1a+Kit+Cd34+類似于preBMPs細(xì)胞的簇,2個Clec12ahi嗜堿性粒細(xì)胞簇(Baso1和Baso2),以及3個Clec12alo嗜堿性粒細(xì)胞簇(Baso3、Baso4和Baso5)。根據(jù)我們的scRNAseq數(shù)據(jù)分析,僅有小部分(11.4%)的Clec12ahi嗜堿性粒細(xì)胞顯示Cd34表達(dá)水平較低,這表明Clec12ahi嗜堿性粒細(xì)胞中的這一小部分可能對應(yīng)于Cd34lo類似于BaP的細(xì)胞。
4、CLEC12Ahi嗜堿性粒細(xì)胞在體外和體內(nèi)分化為CLEC12Alo嗜堿性粒細(xì)胞
對我們的scRNA-seq數(shù)據(jù)進(jìn)行的偽時間軌跡分析和RNA速度分析推斷嗜堿性粒細(xì)胞的分化軌跡,從前BMPs-like細(xì)胞到Clec12ahi嗜堿性粒細(xì)胞,然后到Clec12alo嗜堿性粒細(xì)胞 (圖2g,h)。
為了驗(yàn)證假設(shè),我們追蹤了從骨髓中分離出,并在體外培養(yǎng)2天的c-Kit-嗜堿性粒細(xì)胞的FcεRIαhiCD49blo和FcεRIαloCD49bhi組分的表面CLEC12A和CD9表達(dá)的變化。在培養(yǎng)過程中,大多數(shù)的FcεRIαhiCD49blo嗜堿性粒細(xì)胞的表面表型從CLEC12AhiCD9lo轉(zhuǎn)變?yōu)镃LEC12AloCD9hi(圖3a,左圖)。相反,F(xiàn)cεRIαloCD49bhi嗜堿性粒細(xì)胞即使在培養(yǎng)2天后仍然保持CLEC12AloCD9hi表型(圖3a,右圖)。值得注意的是,F(xiàn)cεRIαloCD49bhi 嗜堿性粒細(xì)胞在2天的培養(yǎng)過程中細(xì)胞總數(shù)減少了約90%,而FcεRIαhiCD49blo 嗜堿性粒細(xì)胞的減少僅約為40%(圖3b,c),這表明前者的壽命較短。當(dāng)我們分別培養(yǎng)骨髓嗜堿性粒細(xì)胞的CLEC12AhiCD9lo和CLEC12AloCD9hi亞群,并追蹤FcεRIα和CD49b表面表達(dá)的變化時,也獲得了相符的結(jié)果。這些結(jié)果強(qiáng)烈表明,CLEC12AhiCD9lo/FcεRIαhiCD49blo 嗜堿性粒細(xì)胞相對不成熟,并且可以分化為CLEC12AloCD9hi/FcεRIαloCD49bhi成熟型嗜堿性粒細(xì)胞,盡管后者的Fcer1a、Mcpt8和Prss34表達(dá)水平低于前者(圖2i)。
接下來,我們檢查了體外實(shí)驗(yàn)的體內(nèi)相關(guān)性。FcεRIαhiCD49blo和FcεRIαloCD49bhi組分分別從CD45.2+小鼠的骨髓細(xì)胞中分類純化并過繼轉(zhuǎn)移到亞致死劑量照射的CD45.1+小鼠(圖3d)。與體外實(shí)驗(yàn)觀察到的結(jié)果一致,大多數(shù)的FcεRIαhiCD49blo 嗜堿性粒細(xì)胞在骨髓中將它們的表面表型從CLEC12AhiCD9lo轉(zhuǎn)變?yōu)镃LEC12AloCD9hi,而大多數(shù)的FcεRIαloCD49bhi 嗜堿性粒細(xì)胞仍保持CLEC12AloCD9hi(圖3d)。
綜上所述,我們得出結(jié)論,骨髓中的CLEC12AhiCD9lo/ FcεRIαhiCD49blo 嗜堿性粒細(xì)胞在表面標(biāo)記物的表達(dá)、細(xì)胞大小和核形狀上顯示出前體like的表型,并在骨髓內(nèi)分化為CLEC12AloCD9hi/ FcεRIαloCD49bhi成熟嗜堿性粒細(xì)胞,這些嗜堿性粒細(xì)胞也存在于外周血和脾臟中。因此,在接下來的實(shí)驗(yàn)中,我們將CLEC12AhiCD9lo/ FcεRIαhiCD49blo和CLEC12AloCD9hi/FcεRIαloCD49bhi 嗜堿性粒細(xì)胞分別稱為前體嗜堿性粒細(xì)胞和成熟嗜堿性粒細(xì)胞。
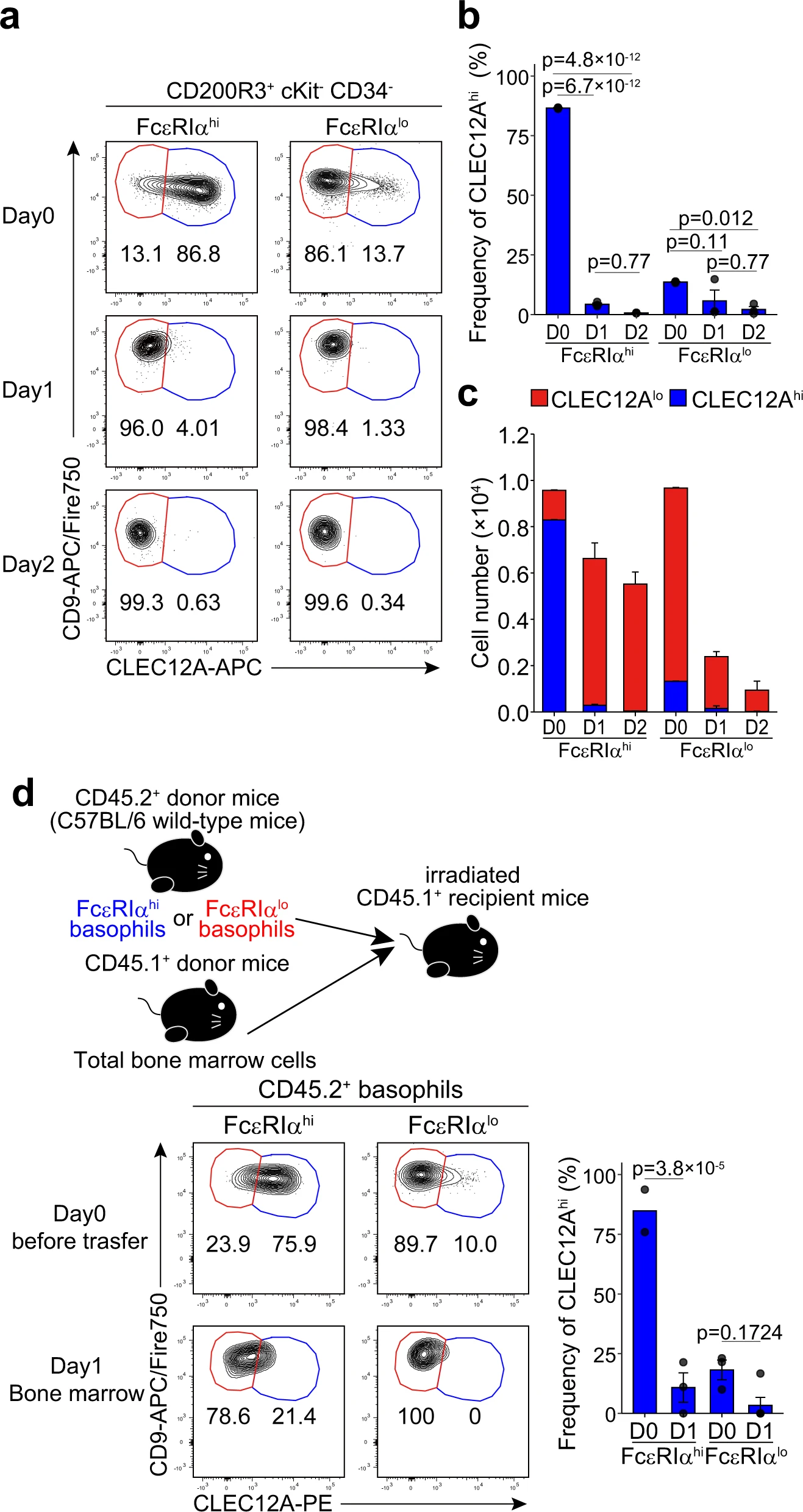
圖3| CLEC12Ahi嗜堿性粒細(xì)胞在體外和體內(nèi)分化為CLEC12Alo嗜堿性粒細(xì)胞
5、前嗜堿性粒細(xì)胞和成熟嗜堿性粒細(xì)胞表現(xiàn)出不同的細(xì)胞增殖和激活特性
為了進(jìn)一步鑒定前嗜堿性粒細(xì)胞(pre-嗜堿性粒細(xì)胞)和成熟嗜堿性粒細(xì)胞(mature 嗜堿性粒細(xì)胞),我們分別從骨髓中分離了CLEC12AhiCD9loCD34-的前嗜堿性粒細(xì)胞以及從骨髓和脾臟分離的CLEC12AloCD9hi的成熟嗜堿性粒細(xì)胞,并利用批量RNA測序分析比較它們的基因表達(dá)譜(圖4a)。作為參考,我們還包括了純化的CD34+ BaPs,它們被定義為單功能嗜堿性粒細(xì)胞前體細(xì)胞。DEGS的系統(tǒng)聚類分析表明,從骨髓和脾臟分離的成熟嗜堿性粒細(xì)胞的基因表達(dá)譜彼此相似,而CD34-的前嗜堿性粒細(xì)胞和成熟嗜堿性粒細(xì)胞的基因表達(dá)譜顯示出明顯差異(圖4a)。值得注意的是,CD34-的前嗜堿性粒細(xì)胞的基因表達(dá)譜與CD34+BaPs的基因表達(dá)譜相似(圖4a),加強(qiáng)了我們之前關(guān)于CD34+BaPs被包括在CLEC12Ahi前嗜堿性粒細(xì)胞群體中的推測。
GO富集分析顯示,與細(xì)胞增殖相關(guān)的GO術(shù)語,如“核分裂”、“染色體分離”和“DNA復(fù)制”,在CLEC12Ahi前嗜堿性粒細(xì)胞中富集,與CLEC12Alo成熟嗜堿性粒細(xì)胞相比(圖4b),單細(xì)胞RNA測序分析結(jié)果顯示,與CLEC12Alo Baso2/3群體相比CLEC12Ahi Baso1群體顯示出較高的S期得分。實(shí)際上,在前嗜堿性粒細(xì)胞中明顯檢測到EdU核苷類似物的攝取,而成熟嗜堿性粒細(xì)胞中未檢測到(圖4d),這表明前嗜堿性粒細(xì)胞的增殖能力。前嗜堿性粒細(xì)胞和成熟嗜堿性粒細(xì)胞在增殖能力上的差異似乎可以解釋在2天培養(yǎng)過程中觀察到的細(xì)胞總數(shù)減少的差異(圖3c)
與前嗜堿性粒細(xì)胞相比,成熟嗜堿性粒細(xì)胞富集了與免疫效應(yīng)功能相關(guān)的GO術(shù)語,如“調(diào)節(jié)免疫效應(yīng)過程”和“正調(diào)節(jié)細(xì)胞因子產(chǎn)生”(圖4c)。當(dāng)受到IgE加抗原的刺激時,CLEC12Alo成熟嗜堿性粒細(xì)胞顯示出比CLEC12Ahi前嗜堿性粒細(xì)胞更高水平的CD63表達(dá)(代表顆粒脫顆粒的指標(biāo))和IL-4產(chǎn)生(圖4e,f),盡管成熟嗜堿性粒細(xì)胞表面FcεRI表達(dá)水平較低(圖2a)。有趣的是,當(dāng)受到IL-3、IL-33和LPS等先天型刺激時,前嗜堿性粒細(xì)胞產(chǎn)生比成熟嗜堿性粒細(xì)胞更高水平的IL-4(圖4g)。此外,批量RNA測序分析發(fā)現(xiàn),在IL-3刺激的前嗜堿性粒細(xì)胞中,其基因表達(dá)譜與IL-3或抗原/IgE刺激的成熟嗜堿性粒細(xì)胞有所不同,包括前者中Il10和Il13的上調(diào)表達(dá)。因此,前嗜堿性粒細(xì)胞似乎對非IgE刺激更具反應(yīng)性,而成熟嗜堿性粒細(xì)胞對IgE/變應(yīng)原刺激更具反應(yīng)性。
根據(jù)細(xì)胞因子環(huán)境的不同即在嗜堿性細(xì)胞產(chǎn)生過程中是否存在IL-3或TSLP ,兩種具有不同表型和功能特性的嗜堿性粒細(xì)胞群體被報(bào)道。當(dāng)受到IL-3和IL-33的刺激時,TSLP誘導(dǎo)的嗜堿性粒細(xì)胞比IL-3誘導(dǎo)的嗜堿性粒細(xì)胞產(chǎn)生更多的IL-4和IL-6。相反,在脫顆粒方面,TSLP誘導(dǎo)的嗜堿性粒細(xì)胞對IgE交聯(lián)的反應(yīng)較差。這種TSLP誘導(dǎo)的嗜堿性粒細(xì)胞的反應(yīng)性似乎與本研究中鑒定的前嗜堿性粒細(xì)胞相似。然而,TSLP誘導(dǎo)的嗜堿性粒細(xì)胞在大小上相對較小,并具有成熟型嗜堿性粒細(xì)胞特有的環(huán)狀核。據(jù)此,我們發(fā)現(xiàn)大多數(shù)TSLP誘導(dǎo)的嗜堿性粒細(xì)胞表現(xiàn)為成熟嗜堿性粒細(xì)胞(FcεRIαloCD49bhi/CLEC12AloCD9hi表型),而不是前嗜堿性粒細(xì)胞。此外,TSLP誘導(dǎo)的成熟嗜堿性粒細(xì)胞顯示出類似于IL-3誘導(dǎo)的成熟嗜堿性粒細(xì)胞的基因表達(dá)譜,而不是前嗜堿性粒細(xì)胞。
綜上所述,TSLP誘導(dǎo)的嗜堿性粒細(xì)胞似乎不對應(yīng)于前嗜堿性粒細(xì)胞,可被歸類為成熟嗜堿性粒細(xì)胞的一個亞群。
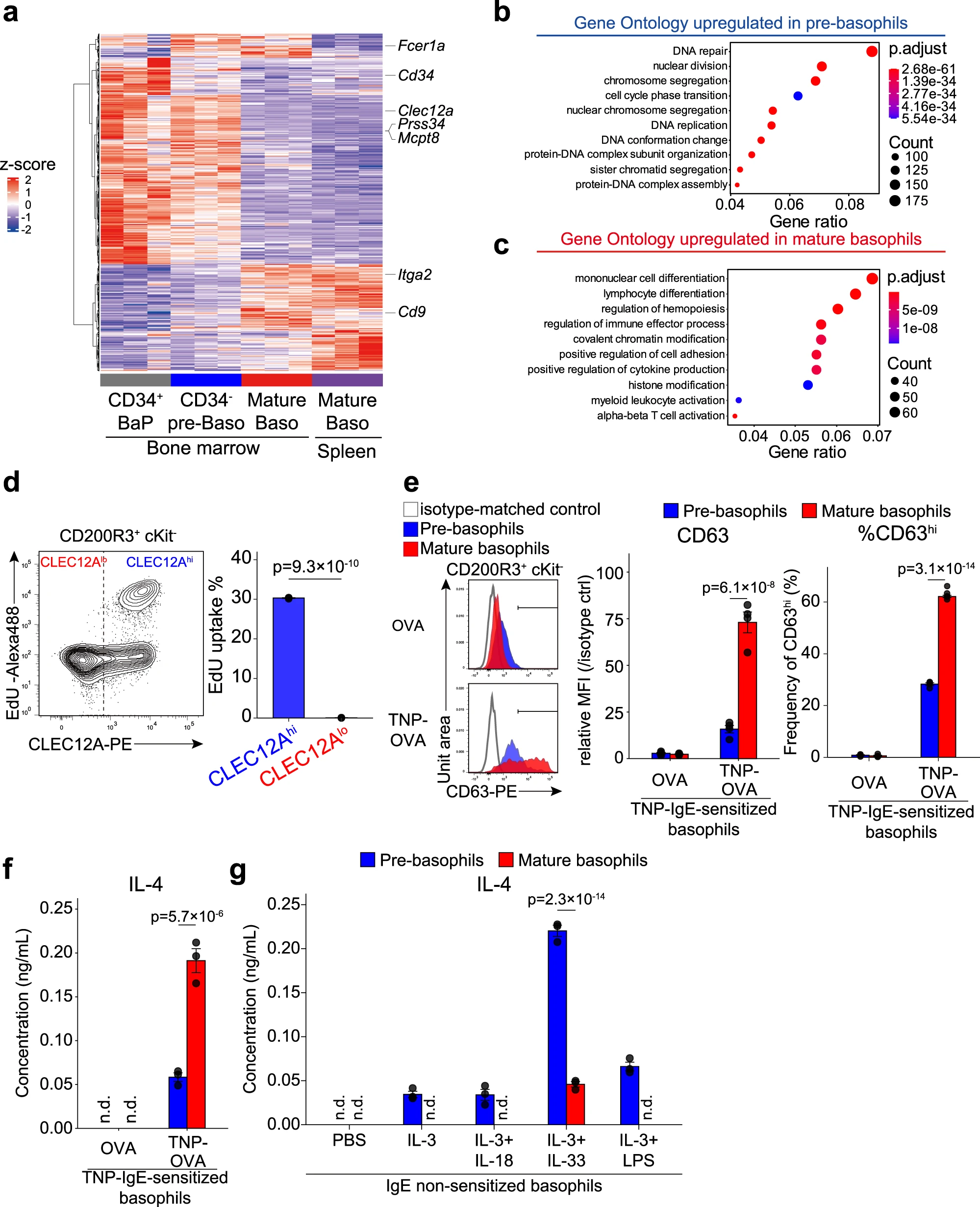
圖4|前嗜堿性粒細(xì)胞和成熟嗜堿性粒細(xì)胞表現(xiàn)出不同的細(xì)胞增殖和激活特性
6、在蠕蟲感染期間,周圍組織中不僅出現(xiàn)成熟的嗜堿性粒細(xì)胞,而且還出現(xiàn)前嗜堿性粒細(xì)胞
在寄生蟲感染期間,嗜堿性粒細(xì)胞常常浸潤到外周組織中。巴西尼普斯壯線蟲(Nb)是一種在人類鉤蟲感染的小鼠模型中被廣泛研究的蠕蟲。它們通過皮膚進(jìn)入宿主動物,遷移到肺部,最終到達(dá)腸道。寄生蟲通過肺部后,嗜堿性粒細(xì)胞在肺部積累,并且對抑制過度肺部炎癥和修復(fù)受損肺組織至關(guān)重要。在Nb感染后的第7天,我們在脾臟、血液甚至肺部除了CLEC12Alo成熟嗜堿性粒細(xì)胞外,還檢測到CLEC12Ahi前嗜堿性粒細(xì)胞(圖5a),這與未感染或PBS注射對照小鼠中前嗜堿性粒細(xì)胞主要存在于骨髓中的觀察結(jié)果相反(圖2a)。從Nb感染的肺部分離的前嗜堿性粒細(xì)胞顯示出比成熟嗜堿性粒細(xì)胞更高的體外EdU摻入(圖5b),這表明前嗜堿性粒細(xì)胞即使在骨髓之外仍保持著較高的增殖能力。此外,在感染的肺部積累的前嗜堿性粒細(xì)胞與成熟嗜堿性粒細(xì)胞的IL-4表達(dá)相當(dāng),這暗示它們可能對Nb感染的Th2免疫起到貢獻(xiàn)作用(圖5c)。
我們以前曾證明在二次Nb感染中,嗜堿性粒細(xì)胞滲入受感染的皮膚,并將蠕蟲困在皮膚內(nèi),以防止感染進(jìn)一步傳播到肺部和腸道。在第二次NB感染的皮損中,除了CLEC12Alo成熟的嗜堿性粒細(xì)胞外,還檢測到了前嗜堿性粒細(xì)胞(圖5d)。從骨髓和第二次感染的皮膚中分離的CD200R3+ cKit-細(xì)胞的單細(xì)胞RNA測序分析鑒定了9個聚類,其中聚類0、1和2對應(yīng)于Cd200r3+ Mcpt8+嗜堿性粒細(xì)胞。在這三個嗜堿性粒細(xì)胞聚類中,我們在兩個器官中都檢測到一個Clec12ahi前嗜堿性粒細(xì)胞聚類和兩個Clec12alo成熟嗜堿性粒細(xì)胞聚類(圖5e,f)。對來自Nb感染和未感染小鼠的嗜堿性粒細(xì)胞的單細(xì)胞RNA測序數(shù)據(jù)進(jìn)行整合分析發(fā)現(xiàn),在Nb感染小鼠的皮膚中檢測到的前嗜堿性粒細(xì)胞的基因表達(dá)譜與未感染小鼠骨髓中的前嗜堿性粒細(xì)胞非常接近,包括Fcer1a,Clec12a,Mcpt8和Prss34的上調(diào)表達(dá)(圖5f)。此外,基因集富集分析(GSEA)和GO富集分析發(fā)現(xiàn),在Nb感染的皮膚中,前嗜堿性粒細(xì)胞富集了與細(xì)胞增殖相關(guān)的基因,包括“染色體分離”(圖5g),這表明前嗜堿性粒細(xì)胞在感染的皮膚中保持著其高增殖能力,在感染的肺部觀察到的結(jié)果也一樣。
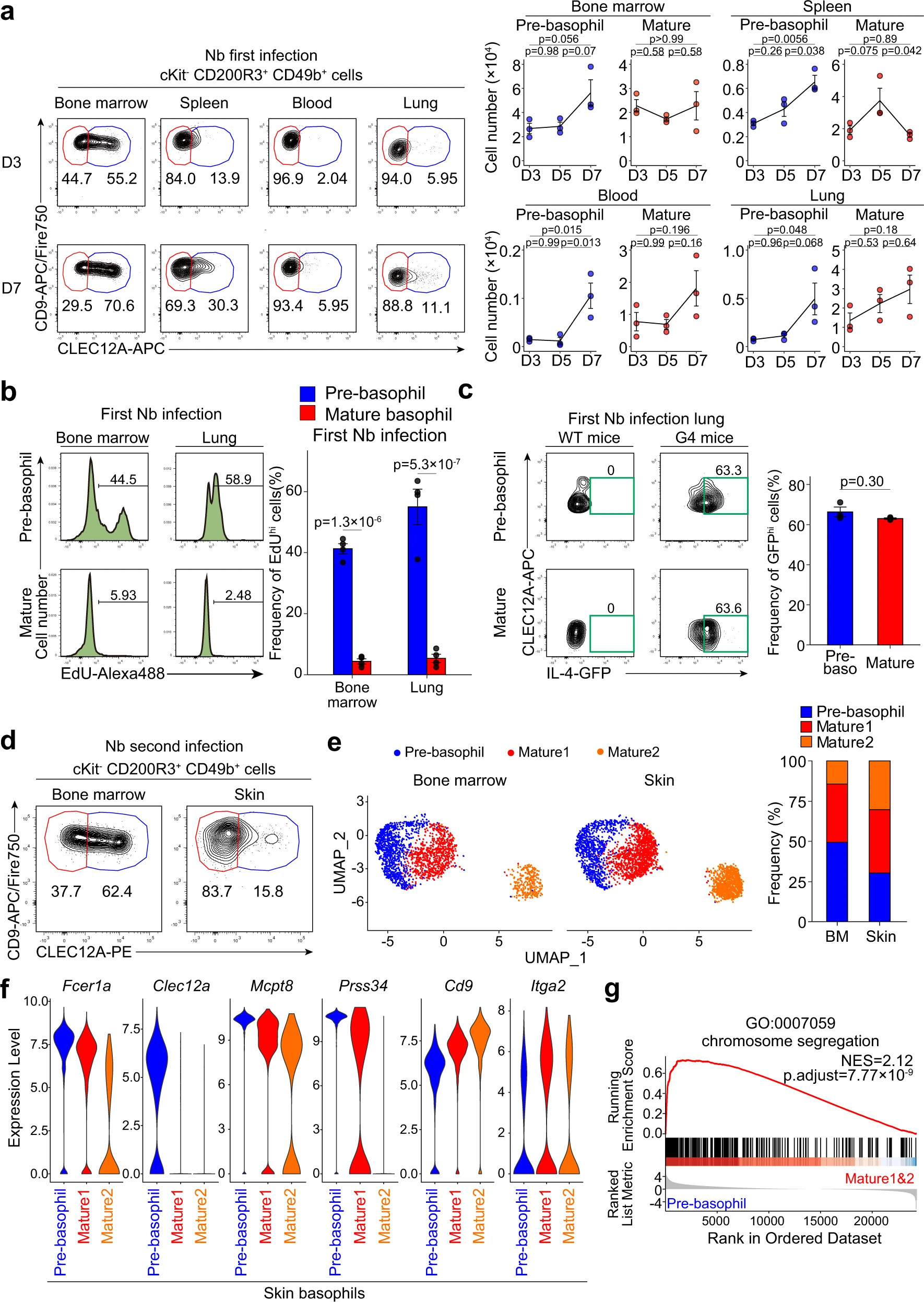
圖5|蠕蟲感染期間外周組織中可檢測到前嗜堿性粒細(xì)胞
7、前嗜堿性粒細(xì)胞中IL-3介導(dǎo)的CXCR4下調(diào)促進(jìn)其從骨髓中排出
接下來,我們試圖闡明在Nb感染期間前嗜堿性粒細(xì)胞在外周組織中可能出現(xiàn)的機(jī)制。
先前已經(jīng)證明Nb感染能夠引起全身性的IL-3上調(diào),因此我們研究了IL-3對骨髓外出現(xiàn)的前嗜堿性粒細(xì)胞的可能貢獻(xiàn)。在小鼠體內(nèi)腹腔注射IL-3復(fù)合物(重組IL-3與單克隆抗IL-3抗體混合)后,外周血中出現(xiàn)CLEC12Ahi的前嗜堿性粒細(xì)胞(圖. 6a)。我們推測IL-3誘導(dǎo)了前嗜堿性粒細(xì)胞從骨髓中外流。為了驗(yàn)證這一可能性,我們對IL-3和PBS對照處理的BMBAs進(jìn)行了批量RNA測序分析,并鑒定了兩者之間的差異表達(dá)基因(DEGs)。我們特別注意到Cxcr4基因的表達(dá)在IL-3刺激下顯著降低(圖. 6b),因?yàn)镃XCR4已被證明是一種重要的趨化因子受體,對免疫細(xì)胞在骨髓中的滯留起作用。骨髓細(xì)胞的單細(xì)胞RNA測序和流式細(xì)胞分析顯示,在轉(zhuǎn)錄和蛋白水平上,CXCR4在前嗜堿性粒細(xì)胞中的表達(dá)高于成熟嗜堿性粒細(xì)胞(圖. 6c),而IL-3刺激能夠降低CLEC12Alo成熟嗜堿性粒細(xì)胞和CLEC12Ahi前嗜堿性粒細(xì)胞表面CXCR4的表達(dá)(圖. 6d)。這些結(jié)果表明,由于CXCR4的高表達(dá),前嗜堿性粒細(xì)胞留在骨髓中,而IL-3介導(dǎo)的CXCR4下調(diào)可能促進(jìn)前嗜堿性粒細(xì)胞從骨髓中外出。根據(jù)這一假設(shè),給小鼠腹腔注射一種CXCR4抑制劑AMD3100,可導(dǎo)致外周血和脾中出現(xiàn)CLEC12Ahi前嗜堿性粒細(xì)胞(圖. 6e)。因此,因此,在骨髓的前嗜堿性粒細(xì)胞中,IL-3介導(dǎo)的CXCR4的下調(diào)至少部分地解釋了前嗜堿性粒細(xì)胞從骨髓中流出,并在Nb感染期間它們出現(xiàn)在肺和皮膚中。
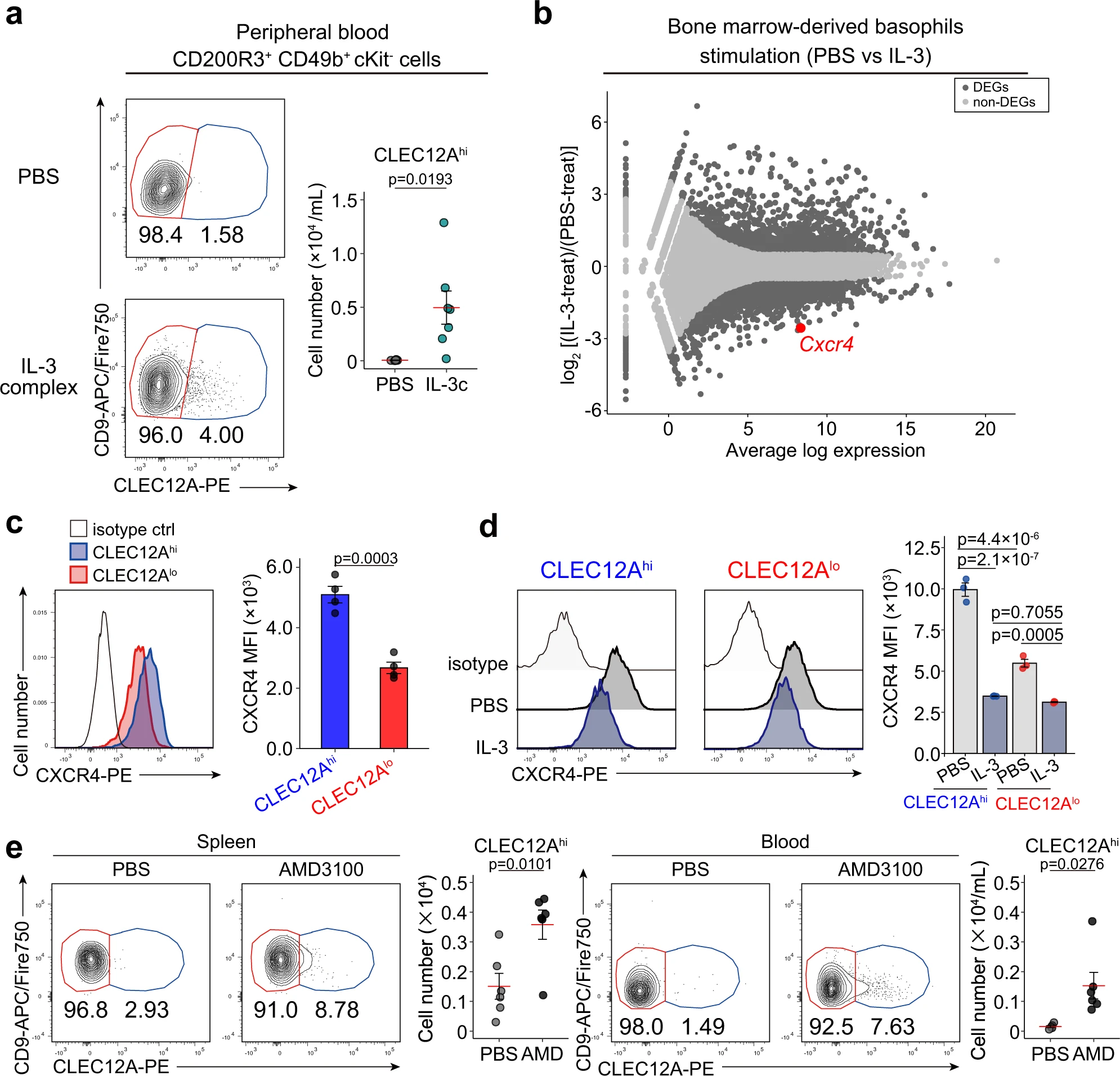
圖6|前嗜堿性粒細(xì)胞中IL-3介導(dǎo)的CXCR4下調(diào)促進(jìn)其從骨髓中排出
參考文獻(xiàn):
Miyake, K., Ito, J., Nakabayashi, J. et al. Single cell transcriptomics clarifies the basophil differentiation trajectory and identifies pre-嗜堿性粒細(xì)胞 upstream of mature 嗜堿性粒細(xì)胞. Nat Commun 14, 2694 (2023).https://doi.org/10.1038/s41467-023-38356-1