






新生物信息技術-EISA:解析轉錄和轉錄后過程對基因表達變化所起的作用
分裂分析(EISA)是一種生物信息技術,通過對RNA-Seq的分析,將轉錄和轉錄后效應分離開來,并將其映射到外顯子和內含子。可測量在不同條件下mRNA和前體RNA水平的變化,可以區分轉錄水平上調控的RNA或在轉錄后水平——miRNA成熟或正在降解。上皮間充質轉化(EMT)是一個備受關注的課題,認識其機制有助于研究腫瘤轉移和藥物敏感性等問題。雖然EMT受許多miRNAs和轉錄因子的調節,然而miRNAs和轉錄因子的功能很難區分,一部分原因是從測序數據集中難以直接識別miRNA靶基因,以及難以確定miRNA效應,本文主要闡述miRNAs參與EMT過程中如何在轉錄組中發揮作用,而運用了一種生物信息技術-EISA(exon-intron split analysis)。(Nucleic Acids Research,IF=11.147 ,Extensive transcriptional responses are co-ordinated by microRNAs as revealed by Exon-Intron Split Analysis (EISA))
研究背景:miR-200c是一個促間質轉化上皮的 microRNA。
一、EISA成功區分轉錄和轉錄后基因調控
![說明: C:\Users\y107\AppData\Roaming\Tencent\Users\3005006148\QiDian\WinTemp\RichOle\H2]IB3(QLO_$HDAA9@PB~~9.png](https://img1.dxycdn.com/2019/0917/713/3368718735652443450-14.jpg)
我們初步比較人乳腺上皮細胞(HMLEs)的內含子豐度的變化與活性和非活性基因的標志的一致性。
圖A:一種通過TGF進入間充質狀態EMT細胞模型(MesHMLE), miR-200C驅動間質向上皮轉化后,前5%的轉錄上調和下調基因 (ΔI),由ChIP-Seq獲得活性染色質(H3K4me3,H3K9-14ac,H3K27c-綠色)和非活性染色質(H3K27me3-紅色)的標記物的相對富集。在一個跨越25個基因滑動窗口中紅色/綠色陰影顯示了測量的均值。(每第5個窗口繪圖)。
圖B:由EISA分析所有組蛋白標記與轉錄水平上的基因表達相關(大ΔI)。相反,活性基因和非活性基因的標記與轉錄后調控的基因(大ΔE-ΔI,小ΔI,圖1B)的相關性小.因此,EISA度量ΔI表明了基因轉錄的影響。基于圖A的ΔE-ΔI的評分。
圖C:對轉染miR-200C的mesHMLE細胞的RNA-seq數據進行EISA分析,以評價miR-200C相對轉錄和轉錄后效應。基于ΔE:ΔI, 轉錄后miR-200 C靶點的測量值負值。
圖D:由ΔE-ΔI、ΔI、ΔE確定基因表達的累積分布(fold change),來預測響應miR-200C的靶位點。
二、miR-200c轉錄后調節EGF信號網絡
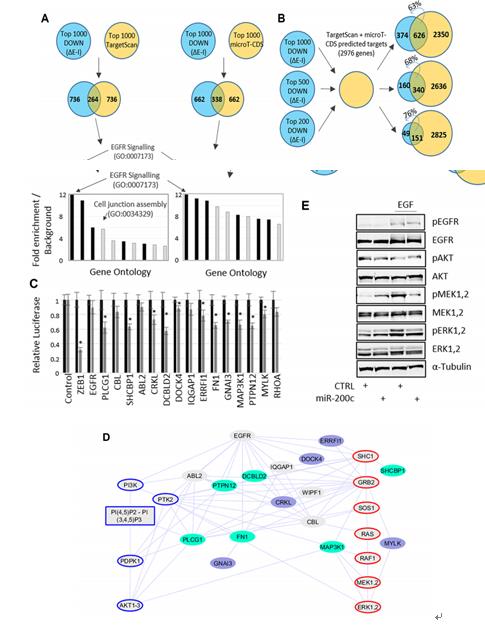
鑒于EISA能有效區分轉錄和轉錄后調節,我們關注在轉錄后下調基因。對miR-200C和預測的靶點進行分析,以及確定在HMLE系統中由miR-200C直接調控的基因和通路。
圖A:通過TargetScan或MicroT-CDS預測miR-200c的靶基因,(ΔE-ΔI,前1000位轉錄后負調控基因)。通過基因本體論研究靶基因之間的關系,發現在EMT中EGFR信號通路的foid enrichment是最高的。
圖B:轉錄后下調基因的大多數是預測的高重疊和預測目標的靶基因。通過the targetscan and microT-CDS algorithms預測,miR200c和miR200c目標反應的1000、500和200個轉錄后下調的基因。
圖C:在EGF信號網絡3’ UTR 熒光素酶報告基因測定miR-200c靶標。
圖D:在EGFR下游的信號網絡中miR-200C的靶基因(藍色陰影)。KEGG pathway顯示Akt和ERK被激活。
圖E:用EGF處理Meshmle細胞,Western Blot檢測在miR-200c表達或不表達的情況下的EGFR、Akt、MEK和ERK(或總蛋白)的磷酸化。
三、在EMT/MET過程中Mir-200 c和TGFβ共同調節的轉錄反應。
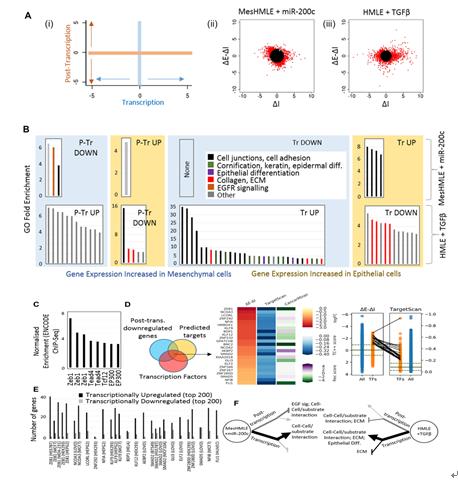
圖A:ΔE-ΔI (轉錄后),ΔI (轉錄)表示EISA定義的基因調控效應. 雖然miRNAs的直接作用是轉錄后產生的,例如miR-200 c對EGF信號的抑制,圖中分別顯示miR-200 c與TGFβ在EMT/MET過程中發生的基因表達變化,其發生ΔE-ΔI (轉錄后)與ΔI (轉錄)。
圖B:雖然已經確定miRNAs和轉錄因子(TGFβ)之間的相互作用,但miRNA研究的一個主要焦點是通過直接作用于靶標來理解其功能。然而,EISA表明,許多對miR-200C的反應是間接的(ΔI)。為了探討這些轉錄變化的意義,基因本體論(分子功能)分析了在MEHMLE細胞對miR-200反應和在HMGLE細胞中對TGF反應的轉錄水平上調和下調基因集。
圖C:在轉錄上調的基因中,例如ZEB 1等,間質促進轉錄因子的結合位點富集在ENCODE ChIP- Seq數據集。每個條形表示單個ChIP- Seq數據集。
圖D:相對于miR-200c(ΔE-ΔI)轉錄后下調的相對程度,顯示了TFs介導下游轉錄變化。轉錄后被miR-200 c下調且在它們的3’端內具有miR-200 c結合位點的TFs。
圖E:TF靶基因是從 ChIP- Seq數據中獲取的,然然后與根據miR-200轉錄上調或下調的基因交叉引用。
四、功能顯著的轉錄調控是miRNAs的一個廣泛特征。
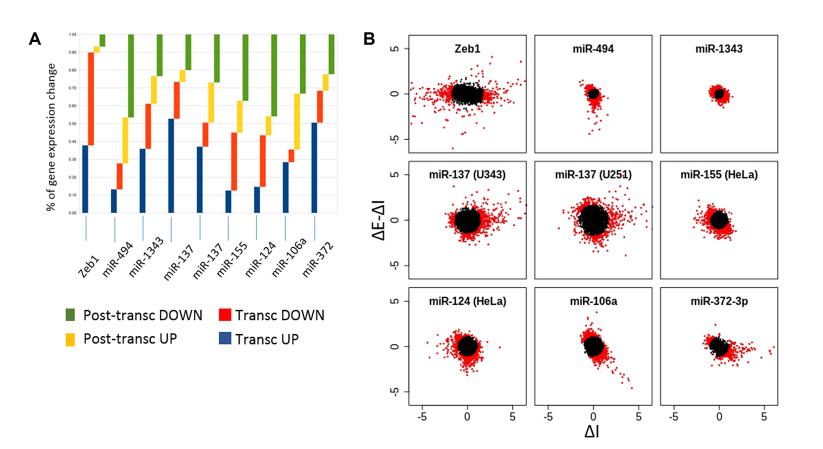

圖A-B:為了研究miRNA干擾中是否與轉錄反應有關,我們對EISA進行了多個數據集的分析,發現在所檢測的8個轉錄反應中,27%到73%的基因是共同的。表達ZEB 1或多個miRNAs的轉錄和轉錄后機制的相對貢獻用散點圖(B)( ΔE-ΔI:ΔI)
圖C:無論這是轉錄上調基因還是下調基因,為每個選定的miRNA進行10個基因 fold enrichment。