






lncRNA Gm10451在iPSC衍生的β樣細胞分化中掩藏的秘密
參考文獻:
Huang Yan., Xu Yang., Lu Yuhua., Zhu Shajun., Guo Yibing., Sun Cheng., Xu Lianchen., Chen Xiaolan., Zhao Yahong., Yu Bin., Yang Yumin., Wang Zhiwei., (2019). lncRNA Gm10451 regulates PTIP to facilitate iPSCs-derived β-like cell differentiation by targeting miR-338-3p as a ceRNA., Biomaterials, 216, 119266. IF:10.273
導語:
iPSC衍生的胰島素生成細胞移植是治療糖尿病比較有前景的一種策略。盡管在過去幾年中已經有許多成熟的體外誘導葡萄糖反應性β細胞的方案,但許多潛在的問題仍有待解決。作為一種關鍵的調節因子,長的非編碼RNA(lncRNA)參與了許多生物過程,包括維持全能性和干細胞分化。在這項研究中,作者發現了一種新的lncRNA Gm10451作為β樣細胞分化的功能調節因子。 Gm10451定位于細胞質,通過靶向miR-338-3p作為競爭性內源RNA(ceRNA)來調節組蛋白H3K4甲基轉移酶復合物PTIP以促進胰島素+ / Nkx6.1 +β樣細胞分化。 miR-338-3p通過靶向PTIP抑制Nkx6.1 +早期β樣細胞分化。移植到鏈脲佐菌素(STZ)小鼠后,β樣細胞中Gm10451的缺失抑制了成熟β細胞的表達,例如胰島素,Nkx6.1和Mafa。雖然小鼠的高血糖癥仍未得到解決,但是該研究為生產更成熟和功能性的iPSC衍生的β樣細胞提供了有效的表觀遺傳靶標。此外,作者推測,將來可用由人類干細胞、生物材料和表觀遺傳修飾產生的胰腺類器官作為新型糖尿病治療的手段。一、差異表達的lncRNA主要參與iPSC衍生的β樣細胞分化 使用3步方案,將小鼠GFP + -iPSC分化成β樣細胞(圖1A)。大約31.63±1.52%(n = 3)的晚期細胞共表達胰島素和Nkx6.1,兩者都是成熟β細胞的標志物(圖1B)。雖然β樣細胞表達胰島素和成熟的β細胞標志物,如Pdx1,Nkx6.1,Mafa,ISL和Glut2,但與成年小鼠胰島相比仍有明顯的差距(圖1C)。與成年小鼠胰島一樣,β樣細胞可響應不同濃度的葡萄糖分泌胰島素(圖1D)。收集4天,14天,21天且時期處在圖1A中步驟3的樣品測序,得到302個差異表達的lncRNAs,2233個差異表達的mRNAs,并做聚類熱圖(圖1E)。得到了6個模式的具有統計學意義的差異表達的lncRNA的表達譜(圖1E)。作者發現共表達網絡中的22個上調的lncRNA和521個mRNA似乎是直接相關的。于是對這22個lncRNA做GO分析,結果顯示,共表達的lncRNA主要富集的生物過程包括轉錄調控,細胞增殖,遷移和凋亡(圖1E)。
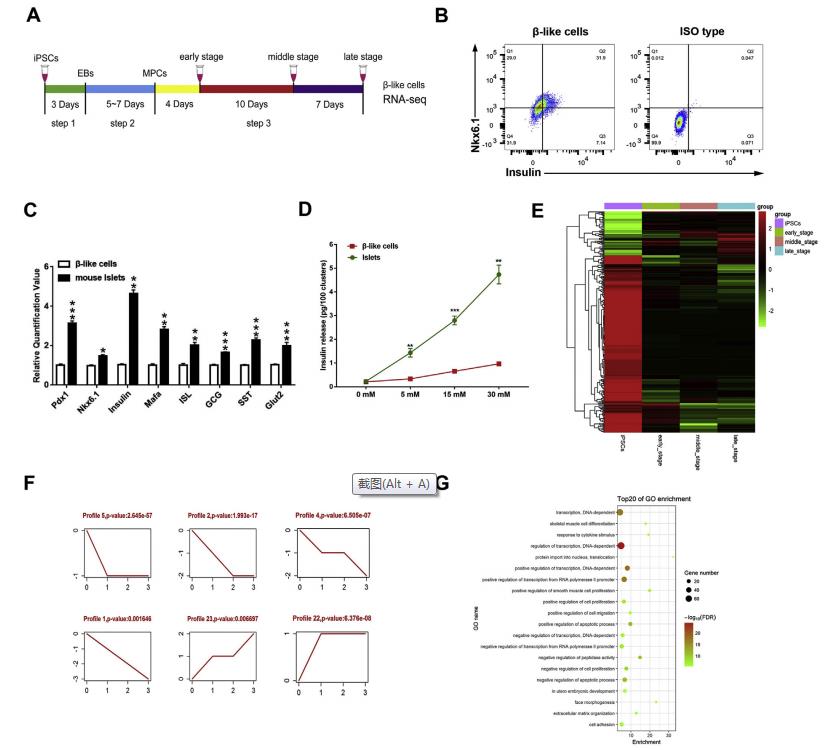
圖1差異表達的lncRNA主要參與iPSC衍生的β樣細胞分化
二、敲除lncRNA Gm10451抑制iPSC-derivedβ-like細胞分化
qRT-PCR驗證測序結果,如圖2所示,lncRNAs Gm10451在分化的早期和中期顯示出明顯的上調表達,選其用于進一步的功能研究(圖2A)。為了確定Gm10451是否是β樣細胞分化的關鍵調節因子,作者構建了siRNA的Gm10451, RT-PCR證實siRNA在早期細胞中實現了約75%的Gm10451敲低(圖2B)。在細胞培養期間,每7天用siRNA轉染細胞以確保Gm10451表達處于相對低的水平,并通過qRT-PCR檢測表達水平(圖2B)。胰腺內分泌祖細胞的所有必需轉錄因子(TF),例如Pdx1,Nkx6.1,Ngn3,Sox9,Pax4 / 6和Gata4 / 6,在Gm10451沉默的細胞中下調(圖2C)。通過免疫熒光染色,我們觀察到在早期分化(步驟3的第6天)中Pdx1,Nkx6.1和Sox9陽性細胞減少(圖2D)。該數據表明, Gm10451可能在β樣細胞的早期分化調節胰腺內分泌祖細胞形成中涉及的關鍵TF的表達中起著重要作用。
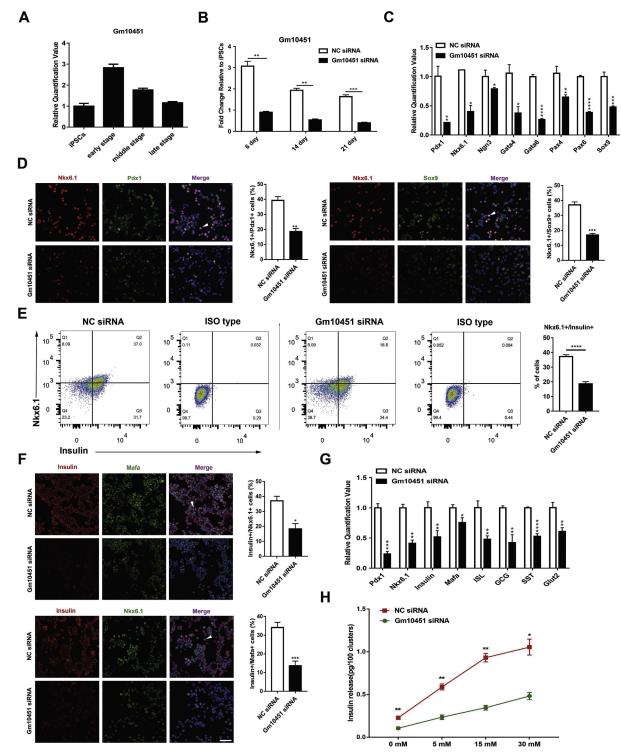
圖2敲除lncRNA Gm10451抑制iPSC-derivedβ-like細胞分化
然后,使用選擇性分化培養基培養Gm10451敲除的細胞15天,收集用于進一步的功能實驗。Gm10451對胰島素+ / Nkx6.1 +β樣細胞群減少的干擾從37.2±1.21%增加到18.6±1.51%(n = 3)(圖2E)。免疫熒光染色也有相似的結果, NC細胞胰島素+和共表達Nkx6.1和Mafa的水平比siRNA組更高(圖2F)。此外,胰島功能標志物mRNA的表達顯著下降,包括胰島素,Glut2,ISL,GCG和SST。并且與對照(NC)細胞相比,成熟β細胞的生物標志物(例如Pdx1,Mafa和Nkx6.1)在Gm10451敲除的細胞中的轉錄水平也顯著下調(圖2G)。 ELISA結果顯示暴露于不同濃度的葡萄糖導致來自對照和Gm10451敲除細胞的胰島素分泌的存在顯著差異。其中,Gm10451敲除的細胞對葡萄糖表現出低敏感性,并且產生和分泌較少的胰島素(圖2H)。
三、新型lncRNA Gm10451的鑒定與表征
細胞分餾分析表明,Gm10451主要定位于細胞質(圖3A)。熒光原位雜交(FISH)的結果也與細胞分餾數據一致(圖3B)。使用RACE技術擴增得到了Gm10451從5’到3’末端的序列(圖2C)。作者用qRT-PCR檢測發現Gm10451的表達水平約為每個細胞50個拷貝。此外,發現新生小鼠的胰腺和肺中富集了Gm10451,這表明Gm10451在胰腺發育中可能發揮功能作用(圖2D)。
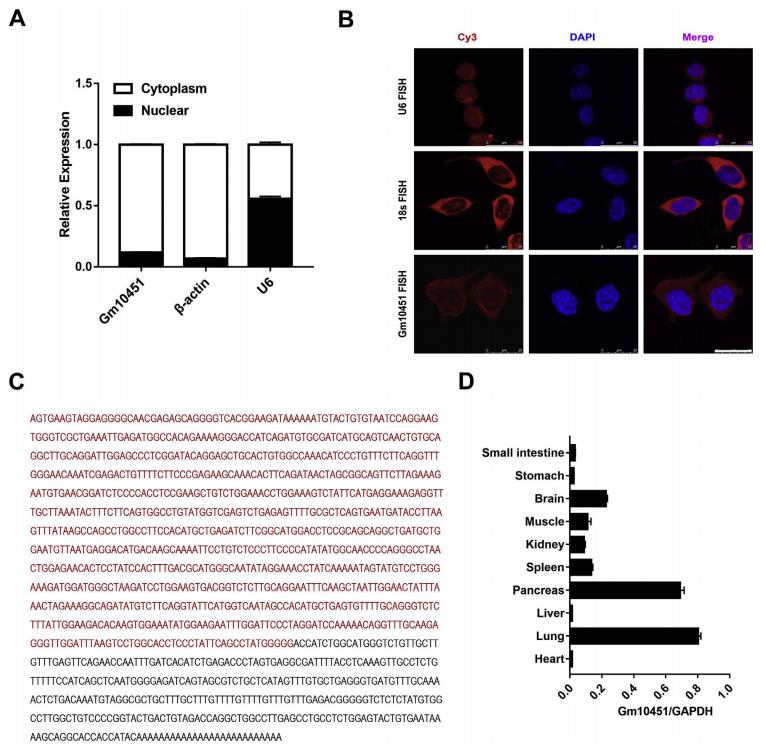
圖3 新型lncRNA Gm10451的鑒定與表征
四、lncRNA Gm10451促進胰島素+ / Nkx6.1 +β樣細胞在體外分化
轉導72小時后,相比于NC,過表達Gm10451細胞中Gm10451基因表達水平上調了3倍多(圖4A)。代表內分泌祖細胞的Nkx6.1 +細胞群分化為β細胞的分化率從27.83±0.76%增加到36.03±1.05%(n = 3)(圖4B)。與NC組相比,胰腺β-細胞分化的關鍵TF顯著上調(圖4C)。比較Gm10451過表達和對照細胞中胰島關鍵標志物的表達,發現兩組之間胰島素Glut2,ISL,Pdx1和Mafa的轉錄水平存在顯著差異(圖4D)。Gm10451過表達促使β樣細胞響應低濃度的葡萄糖(5mM)迅速分泌胰島素(圖4E),胰島素分泌在葡萄糖濃度為15-30 mM時達到穩定水平(圖4E)。胰島素+ / Nkx6.1 +β樣細胞的數量從31.3±1.19%顯著增加至49.83±2.02%(n = 3),進一步表明Gm10451可作為成熟胰島素分泌細胞分化的調節劑(圖4F)。
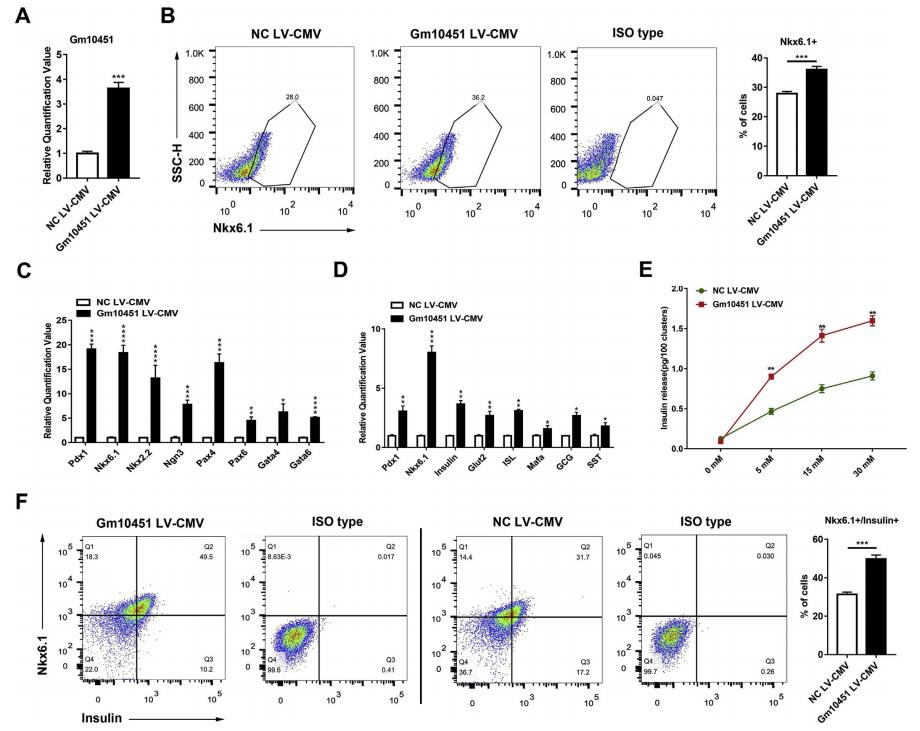
圖4 lncRNA Gm10451促進胰島素+ / Nkx6.1 +β樣細胞在體外分化
五、 mir - 338 - 3 - p調節早期β樣 PTIP表達和抑制細胞分化
為了確定哪種miRNA可能參與調節PTIP,利用免疫印跡檢測PTIP水平,結果顯示miR-338-3p顯著下調PTIP水平(圖5A)。熒光素酶測定系統驗證miR-338-3p可以抑制PTIP的翻譯(圖5B)。熒光素酶報告基因測定顯示PTIP的野生型3'-UTR僅在miR-338-3p存在時翻譯水平較低(圖5C)。突變的3'-UTR對miR-338-3p沒有顯示出統計學上的顯著改變(圖5C)。Nkx6.1 +細胞群分化率從31.06±1.00%顯著降低至23.13±1.03%(n = 3)(圖5D)。除了PTIP之外,胰腺內分泌祖細胞的幾個關鍵轉錄因子,例如Pdx1,Nkx6.1,Ngn3,Pax6和Gata6,都顯著下調(圖5E)。 免疫熒光染色也證明miR-338-3p過表達在分化早期降低了PTIP,Nkx6.1和Ngn3陽性細胞數(圖5F)。與陰性對照(21.6±1.51%,n = 3)相比,miR-338-3p敲低部分逆轉了Gm10451表達不足對損傷分化的影響(圖5G)。
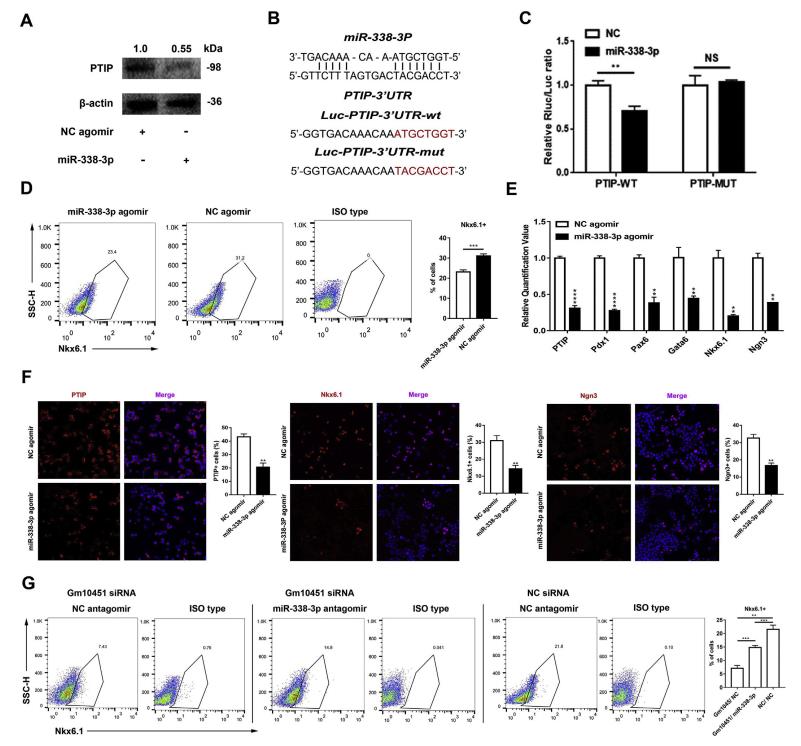
圖5 mir - 338 - 3 - p調節早期β樣 PTIP表達和抑制細胞分化
六、Gm10451直接結合miR-338-3p并通過miR-338-3p/PTIP軸調節β樣細胞分化
在線預測Gm10451和miR-338-3p的結合位點,從484到495(圖.6A)。構建Gm10451(LucGm10451-wt)和突變體(Luc-Gm10451-mut)的熒光素酶載體(圖6A)。熒光素酶測定顯示,miR-338-3p可以抑制Gm10451的熒光素酶活性,但它對突變的Gm10451的影響較小,這表明Gm10451可以使用預測的結合位點與miR-338 3p相互作用(圖6B)。敲除Gm10451會導致PTIP表達下調(圖6C)。Gm10451過表達導致PTIP表達上調(圖6D)。Gm10451中和了miR-338-3p對PTIP表達的抑制作用(圖6E)。拯救實驗表明,過表達的Gm10451可逆轉PTIP-siRNA對胰島素+細胞的抑制作用(圖6F)。qRT-PCR結果顯示,過表達的Gm10451也拯救了PTIP敲除細胞中重要基因的轉錄水平(圖6G),表明Gm10451可通過miR-338-3p / PTIP軸調節iPSC衍生的β樣細胞分化。
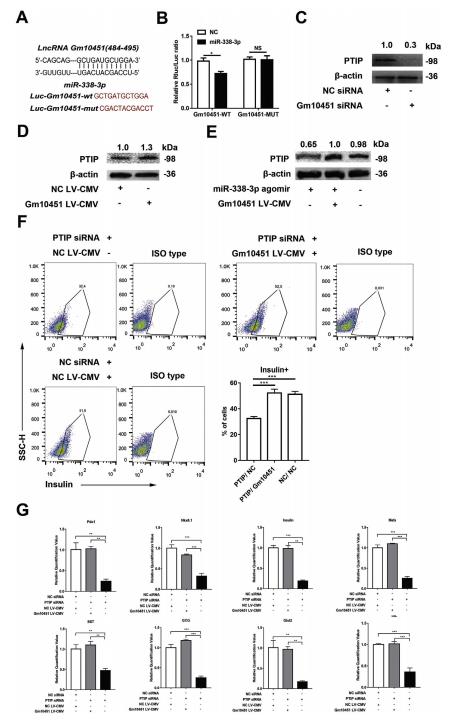
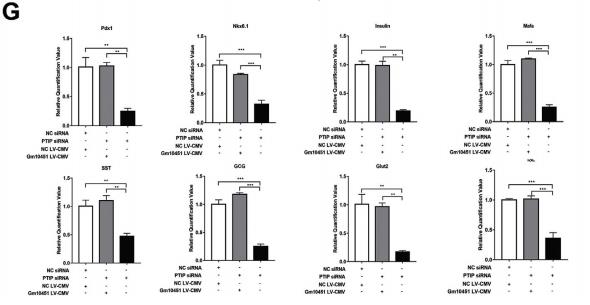
圖6 Gm10451直接結合miR-338-3p并通過miR-338-3p / PTIP軸調節β樣細胞分化
七、胰腺內分泌祖細胞中PTIP敲除后,全基因組H3K4me3減少
用siRNA沉默MPC中的PTIP,導致H3K4me3的表達減少(圖7A)。H3K4me3富集變化的分布餅圖顯示大多數位于基因間或內含子區域內(圖7B)。火山圖顯示,中101個基因與H3K4me3相關性在NC組中比PTIP-siRNA組更多(圖7C)。 GO分析顯示,差異表達的基因富含多個生物過程,包括細胞生物學和生物體相關過程,這些過程在β細胞分化和形成中很重要(圖7D),qRT-PCR驗證了Mrpl15,Smyd3,Cnot2,Nrf1,Nr4a1和Igf1r,CHIP-seq結果顯示它們H3K4me3的富集水平都較低(圖7E)。
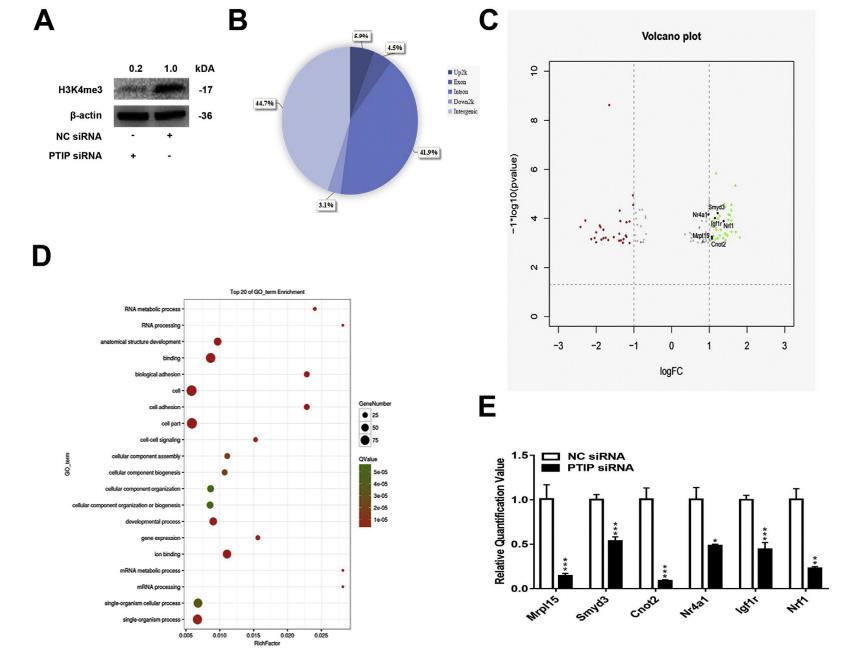
圖7胰腺內分泌祖細胞中PTIP敲除后,全基因組H3K4me3減少
八、 Gm10451沉默的β樣細胞不能逆轉鏈脲佐菌素(STZ)誘導的糖尿病小鼠的高血糖癥
為了比較來自NC和Gm10451-siRNA組的β樣細胞在糖尿病小鼠中的功能,將去細胞化的大鼠胰腺皮下移植到小鼠的背部(圖8A)。每組大約1×106個β樣細胞被植入,在移植后監測空腹血糖水平59天(圖8B)。在移植后29天,種植的NC β樣細胞小鼠血糖水平恢復到基線,但注射Gm10451-siRNAβ樣細胞不能完全逆轉高血糖癥。最后,在移植物取出后5天內NC β樣細胞小鼠血糖恢復至高血糖水平(圖8B)。移植后59天取出移植物,免疫熒光染色顯示與Gm10451-siRNA組相比,NC組含有更多的胰島素+細胞,并且Nkx6.1和Mafa顯著高表達(圖8C)。因此,結果表明Gm10451在體內和體外可以通過靶向miR-338-3p作為ceRNA來調節PTIP以促進iPSC衍生的β樣細胞分化(圖8D)。
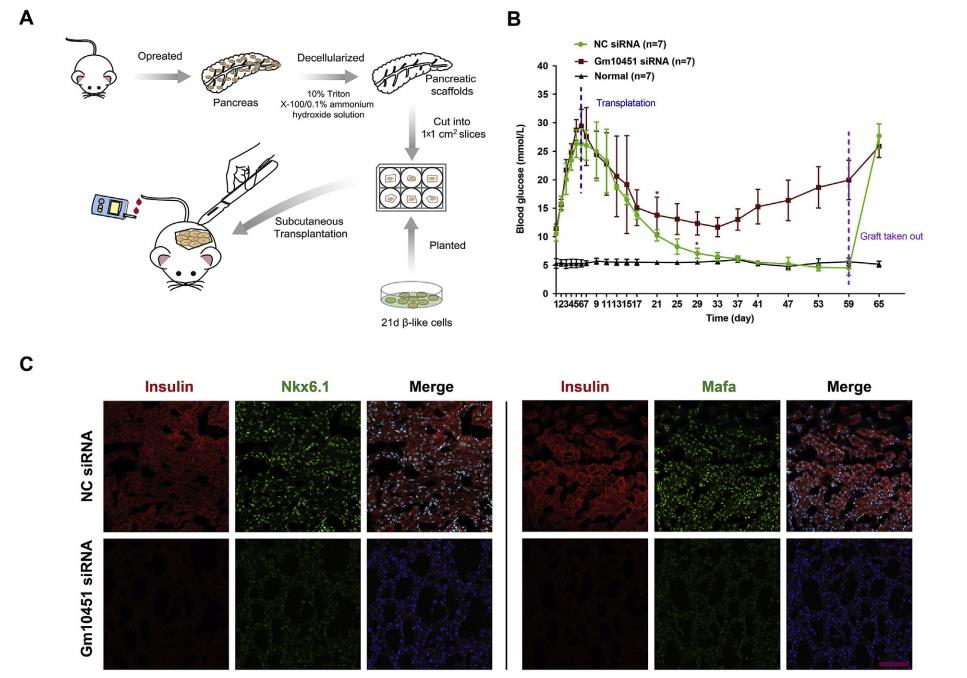
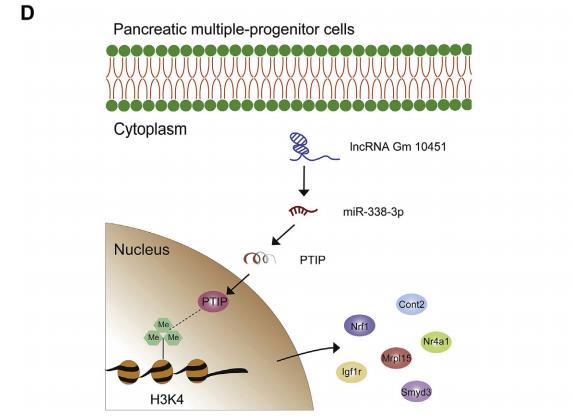
圖8 Gm10451沉默的β樣細胞不能逆轉鏈脲佐菌素(STZ)誘導的糖尿病小鼠的高血糖癥