






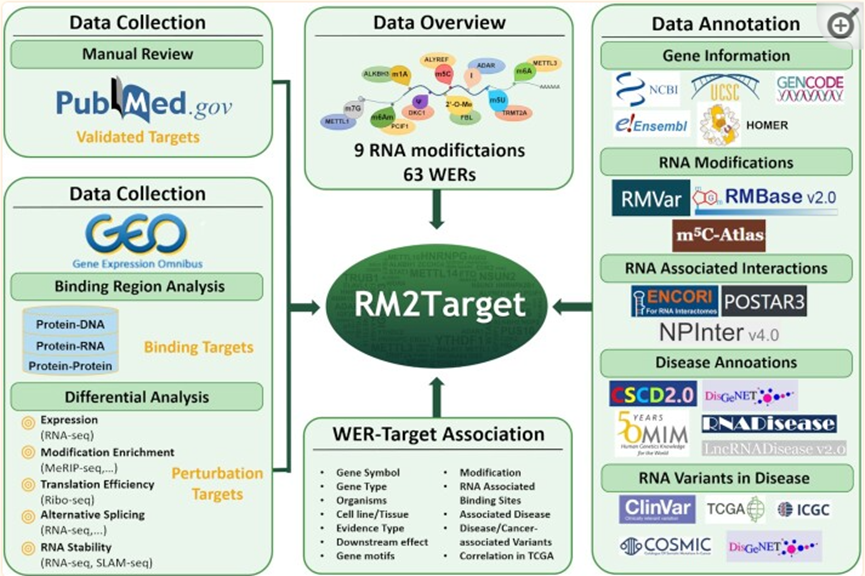
RM2Target:RNA修飾數(shù)據(jù)庫
RNA修飾在調(diào)節(jié)RNA代謝中發(fā)揮著關(guān)鍵作用,如基因轉(zhuǎn)錄、RNA穩(wěn)定性、RNA剪接、核定位和翻譯。RNA修飾是由三類不同的蛋白質(zhì)動態(tài)介導(dǎo)的:寫入者、擦除者和讀取器(WER)。寫入者可以調(diào)節(jié)RNA修飾的沉積;例如,METTL3和NSUN2可以分別將N6-甲基腺苷(m6A)和5-甲基胞嘧啶(m5C)寫入其靶基因。擦除者去除靶基因的RNA修飾;例如,FTO是m6A的擦除器,而ALKBH3是N1甲基腺苷(m1A)的擦除器。讀取器通過識別靶基因中的RNA修飾位點來發(fā)揮其功能;例如YTHDF1/2/3是m6A(6)的讀取器并且ALYREF是m5C的讀取器。WER的失調(diào)已被證明與各種疾病有關(guān),包括癌癥、心血管疾病和神經(jīng)系統(tǒng)疾病。相同的WER在不同的條件下可能具有不同的功能。例如,METTL3在大多數(shù)癌癥類型中起致癌作用,但據(jù)報道,它在某些癌癥類型中也具有腫瘤抑制功能,如腎癌。這主要是因為WER的擾動可能在不同的條件下選擇性地影響不同的靶基因集。總之,識別WER-靶標(biāo)關(guān)聯(lián)對于研究RNA修飾在各種生理和病理條件下的功能和調(diào)節(jié)機制尤為重要。然而,目前還沒有公共存儲庫來承載不同RNA修飾的WER-靶標(biāo)關(guān)聯(lián)。
得益于實驗和高通量測序技術(shù)的發(fā)展,可以為探索WER與靶基因之間的關(guān)系提供更多的證據(jù)。例如,免疫沉淀和某些下一代測序技術(shù),如RIP-seq和CLIP-seq,可以直接有效地闡明WER蛋白與靶RNA之間的結(jié)合關(guān)系。微擾技術(shù),如小干擾RNA(siRNA)和CRISPR/Cas9系統(tǒng)以及高通量測序,可用于系統(tǒng)評估WERs對特定基因RNA代謝的微擾效應(yīng)。在過去的兩年里,與RNA修飾相關(guān)的研究激增。隨著更多WER的發(fā)現(xiàn),靶基因的數(shù)量也有所增加,因此需要更新m6A2Target。因此研究團隊升級了數(shù)據(jù)庫并取名為RM2Target (http://rm2target.canceromics.org/)。來自兩種生物體和九種類型的RNA修飾的63個WER的靶標(biāo)關(guān)聯(lián),包括N6甲基腺苷(m6A)、N6,2'-O-二甲基腺苷(m6Am)、N1甲基腺苷(m1A)、假尿苷(ψ)、5-甲基尿苷(m5U)、5-甲基胞嘧啶(m5C)、7-甲基鳥苷(m7G)、2'-O-甲基化和A-to-I RNA編輯。RM2Target含有包含1619653個WER,為了讓用戶進一步研究WER和靶基因的功能,并研究RNA修飾與疾病之間的關(guān)系,RM2Target提供了基本的基因信息和豐富的注釋,包括RNA修飾、RNA-RNA/RNA-蛋白質(zhì)相互作用、TCGA癌癥類型中WER及其靶基因之間的表達相關(guān)性,以及RNA與疾病的關(guān)聯(lián)。
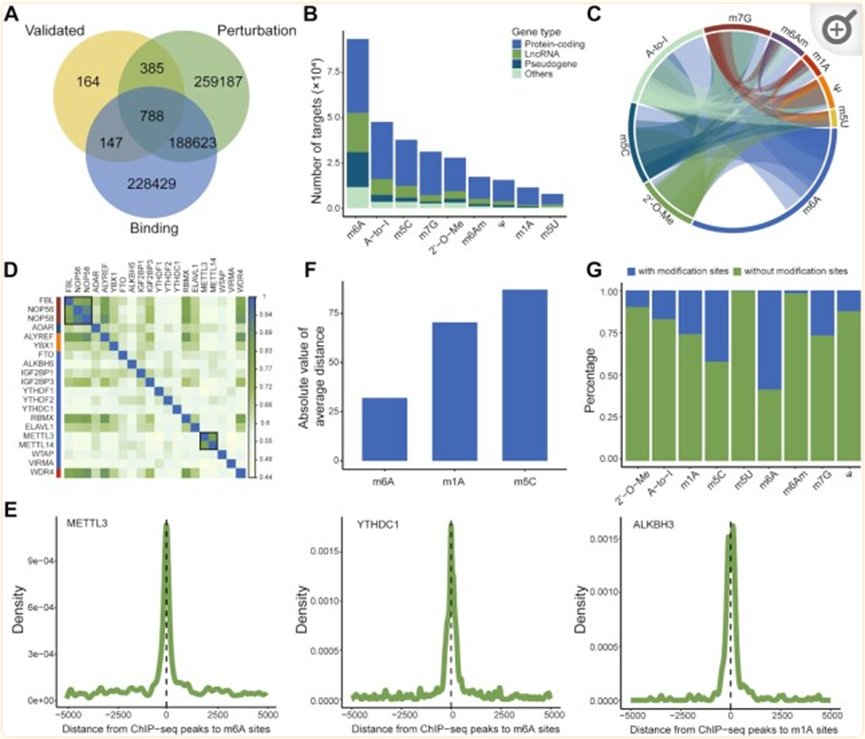
三種證據(jù)類型的WER-靶標(biāo)關(guān)聯(lián)存儲在RM2Target中:(i)“已驗證”:分別使用蛋白質(zhì)印跡、RT-qPCR、RNA穩(wěn)定性測定、RIP測定、熒光素酶報告基因測定等方法,通過體內(nèi)或體外實驗驗證人類和小鼠的1530和584個WER-靶點關(guān)聯(lián)。(ii)“結(jié)合”:通過對人類和小鼠的高通量分析,分別預(yù)測了461746和61395 WER-靶標(biāo)與結(jié)合證據(jù)的關(guān)聯(lián)。其中,人類有451016個WER-靶標(biāo)與蛋白質(zhì)-RNA相互作用的關(guān)聯(lián),小鼠有55004個。在人類中總共記錄了3100個WER-靶標(biāo)與蛋白質(zhì)-蛋白質(zhì)相互作用的關(guān)聯(lián)。(iii)“擾動”:分別從人類和小鼠的WER擾動中推斷出646 539和447 859 WER-靶標(biāo)關(guān)聯(lián)。先前的研究表明,RNA修飾在某些基本的生物過程中發(fā)揮著至關(guān)重要的作用,如mRNA的穩(wěn)定性、剪接和翻譯。因此,RM2Target在五個水平上進行了差異分析,即表達、修飾、翻譯、穩(wěn)定性和選擇性剪接。對于人類,有339604個WER-靶標(biāo)與RNA水平變化相關(guān),60176個與修飾水平變化有關(guān),117866個與翻譯效率變化有關(guān),27700個與mRNA穩(wěn)定性變化有關(guān),101193個與差異選擇性剪接事件有關(guān)。
RM2Target揭示WER與靶基因之間的復(fù)雜調(diào)控網(wǎng)絡(luò)
不同WER之間的串?dāng)_同一RNA修飾的寫入者、擦除者和讀取器經(jīng)常合作控制細胞表型。越來越多的證據(jù)表明,不同的RNA修飾之間存在頻繁的串?dāng)_。例如,NSUN2介導(dǎo)的m5C修飾可以促進METTL3介導(dǎo)的m6A修飾,反之亦然。m6A修飾可能通過阻斷ADAR和靶基因的結(jié)合來抑制A-to-I編輯水平。我們系統(tǒng)地分析了在RM2Target中收集的數(shù)據(jù),以探索不同RNA修飾的不同WER之間的串?dāng)_。觀察到來自不同修飾的WER的靶基因之間的密集交聯(lián)。正如預(yù)期的那樣,m6A寫入者METTL3和METTL14的靶基因具有高度重疊,與METTL3和METTL14通常作為異二聚體復(fù)合物起作用的事實一致。此外,FBL、NOP56和NOP58,2′-O-甲基化的作者,被證明是盒小核仁核糖核蛋白復(fù)合物(snoRNPs)的核心蛋白,也共享相似的靶基因集。
RNA修飾和基因轉(zhuǎn)錄之間的串?dāng)_RNA修飾的沉積通常被認(rèn)為是一個共轉(zhuǎn)錄過程。RNA修飾也可以調(diào)節(jié)基因轉(zhuǎn)錄。通過對WER的ChIP-seq數(shù)據(jù)和RNA修飾位點的峰位點進行綜合分析,我們發(fā)現(xiàn)m6A-WER的ChIP-seq峰位點,如METTL3、YTHDC1,靠近相應(yīng)的m6A位點,m1A-WER峰位點,例如ALKBH3,接近相應(yīng)的m1A位點,表明這些RNA修飾和基因轉(zhuǎn)錄之間存在強烈的串?dāng)_。我們計算了WER的ChIP-seq峰中心和最近的RNA修飾位點之間的平均距離,并認(rèn)為該平均距離表明了RNA修飾和基因轉(zhuǎn)錄之間的串?dāng)_。因此,我們觀察到m6A修飾主要涉及串?dāng)_,m5C最少涉及串?dāng)_。
與RNA修飾無關(guān)的WER-靶標(biāo)關(guān)聯(lián)最近的研究表明,一些已知的RNA修飾的WER具有與修飾無關(guān)的功能。例如,m6A寫入器METTL3可以在沒有m6A修飾的情況下促進靶基因PAPBC1的翻譯。m6A寫入物METTL16發(fā)揮m6A非依賴性功能以促進翻譯和腫瘤發(fā)生。我們使用RM2Target數(shù)據(jù)系統(tǒng)地研究了RNA修飾依賴性和非依賴性WER-靶標(biāo)關(guān)聯(lián)。我們發(fā)現(xiàn),只有30%的靶基因具有來自公共數(shù)據(jù)庫(如RMbase和RMVar)的相應(yīng)RNA修飾位點,這表明WERs可能具有RNA修飾無關(guān)的功能。然而,目前檢測RNA修飾的方法可能由于各種原因錯過了許多RNA修飾位點,這一可能性不容忽視。
