






scRNA-seq和scATAC-seq整合分析發高分——揭示iNKT細胞發育軌跡
與傳統的αβT細胞不同,固有自然殺傷T細胞(iNKT細胞)在胸腺中完成其終末分化為功能性的iNKT1/2/17細胞。然而,指導iNKT亞群分化的潛在分子程序仍然不清楚。在這里,作者利用單細胞RNA測序(scRNA-seq)和單細胞轉座可及染色質測序(scATAC-seq)對四個胸腺iNKT發育階段的超過17,000個iNKT細胞的轉錄組和超過39,000個iNKT細胞的染色質可及狀態進行了分析,以定義它們的發育軌跡。研究發現了iNKT前體和不同iNKT亞群的新特征,并表明iNKT2和iNKT17細胞系的發育可能在第0階段(ST0)早期就通過兩個不同的程序發生了,而iNKT1的發育可能發生在ST0之后。iNKT1和iNKT2細胞都表現出廣泛的表型和功能多樣性,而iNKT17細胞相對均一。此外,發現一個新的轉錄因子Cbfβ在iNKT前體中高表達,其表達軌跡與其他已知的iNKT細胞發育轉錄因子Zbtb16和Egr2相似,并且可以指導iNKT細胞命運并推動它們的效應器表型分化。通過條件性敲除Cbfβ,阻斷了早期iNKT細胞的發育,并導致iNKT1/2/17細胞分化嚴重受損。總的來說,本研究揭示了不同的iNKT發育程序以及它們的細胞多樣性,并確定了一個新的轉錄因子Cbfβ作為早期iNKT細胞發育的關鍵調節因子。本文于2023年6月發表在《Cell Discovery》IF: 33.5期刊上。
技術路線

主要實驗結果:
1、通過scRNA-seq和scATAC-seq對胸腺iNKT細胞在連續發育階段進行聚類
為揭示iNKT細胞的發育景觀,采用熒光激活細胞分選(FACS)技術從連續發育階段中收集胸腺iNKT細胞,以進行scRNA-seq和scATAC-seq實驗(圖1a,b)。鑒于在C57BL/6小鼠中胸腺ST0(CD24+) iNKT細胞的稀缺性,利用Vα14-Jα18轉基因小鼠(也稱為rec-Vα14Tg)進行ST0 iNKT細胞分析,其TCR基因座與內源性TCR基因座非常相似,但具有豐富的ST0 iNKT細胞。如預期的,發現在rec-Vα14Tg和C57BL/6小鼠的ST0 iNKT細胞中,scATAC-seq譜的相似性很高。因此,進一步使用rec-Vα14Tg小鼠進行ST0 iNKT細胞的scRNA-seq分析,并使用C57BL/6小鼠進行ST1(CD24?CD44loNK1.1?)、ST2(CD24?CD44hiNK1.1?)和ST3(CD24?CD44hiNK1.1+) iNKT細胞的分析(圖1a–c)。
經過質量控制過濾和排除細胞異常值后,共保留了17,944個高質量的單一胸腺iNKT細胞,總共表達了13,578個基因,用于隨后的scRNA-seq分析。在聚合的iNKT細胞(ST0–ST3)中鑒定16個簇,每個簇的細胞數量從255個到2817個不等(圖1d,e)。每個簇中最顯著差異表達的基因(DEGs)顯示在熱圖中(圖1f)和小提琴圖中(圖1g)。在這些簇中,有四個簇(C2,C4,C9和C14)來自ST0,八個簇來自ST1(C3,C5,C6,C7,C10,C11,C15和C16),九個簇來自ST2(C1,C3,C5,C6,C7,C10,C11,C13和C15),以及四個簇來自ST3(C1,C5,C8和C12)(圖1d)。ST0 iNKT細胞明顯與其他iNKT階段分離,而來自ST1和ST2 iNKT細胞的簇有適度的重疊,而來自ST3 iNKT細胞的簇與來自ST2 iNKT細胞的簇緊密相鄰(圖1d)。此外,相關性分析表明,同一階段內的不同簇呈相對相似的轉錄組模式。
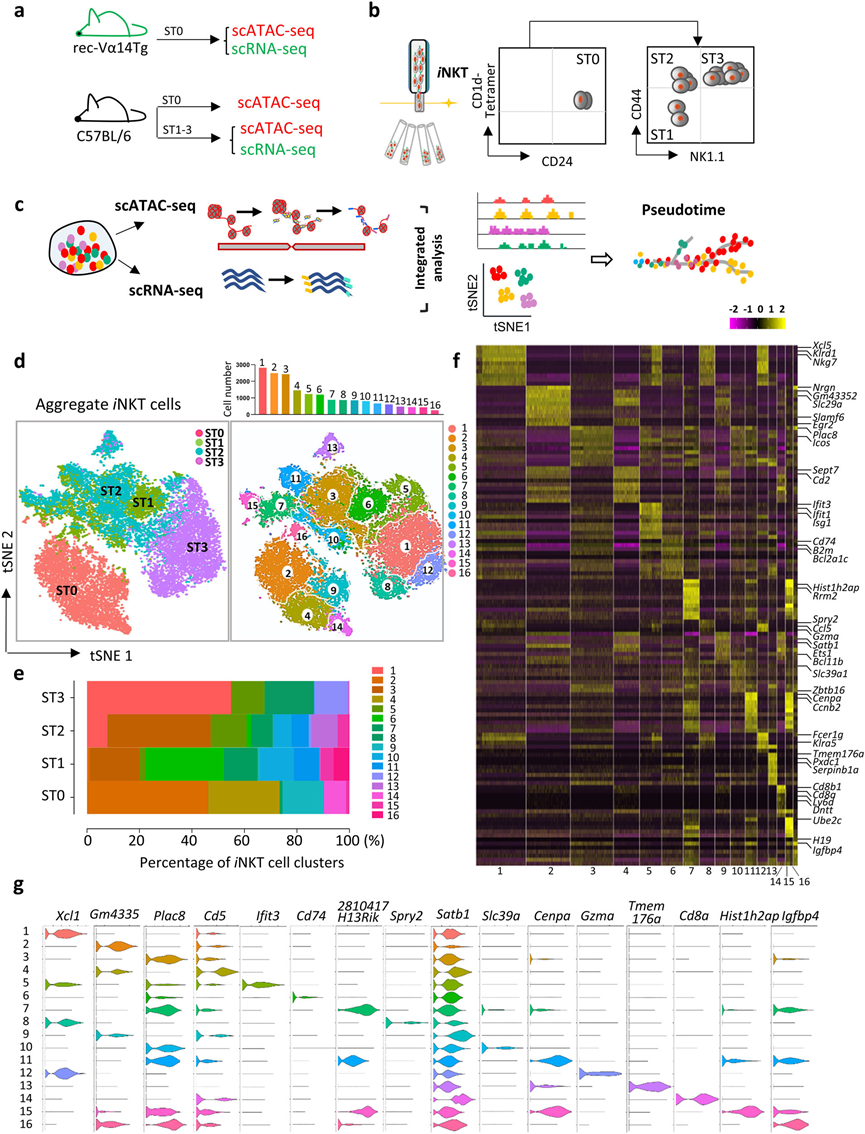
圖1 小鼠胸腺iNKT細胞的多樣性
細胞分化伴隨著由順式調控元件控制的基因的表達,這些元件必須處于開放狀態才能正常發揮功能。因此,對與scRNA-seq分析中相同發育階段的胸腺iNKT細胞進行了scATAC-seq分析,并繪制了單個iNKT細胞的染色質可及性景觀。在與scRNA-seq數據整合后,這十六個iNKT細胞簇很容易識別(圖2a)。由于scRNA-seq允許根據轉錄組推測當前細胞狀態,作者根據其特征轉錄組和細胞因子轉錄表達,將每個簇(C1–C16)分配到已發表的iNKT1、iNKT2和iNKT17功能亞群中。結果發現來自ST1和ST2的C3、C6、C7、C10、C11、C15和C16簇被分類為iNKT2亞群;來自ST3主要的C1、C5、C8和C12簇被分類為iNKT1(圖2b);而來自ST2的獨特iNKT C13被分配到iNKT17(圖2b)。ST0簇(C2、C4、C9和C14)在任何iNKT亞群中都不突出,因為它們沒有表現出強烈的效應特征(圖2b)。總體而言,通過整合轉錄組和表觀遺傳學特征,繪制了胸腺iNKT細胞的動態轉錄組和染色質景觀,揭示了iNKT1和iNKT2細胞的異質性以及iNKT17細胞的相對同質性。
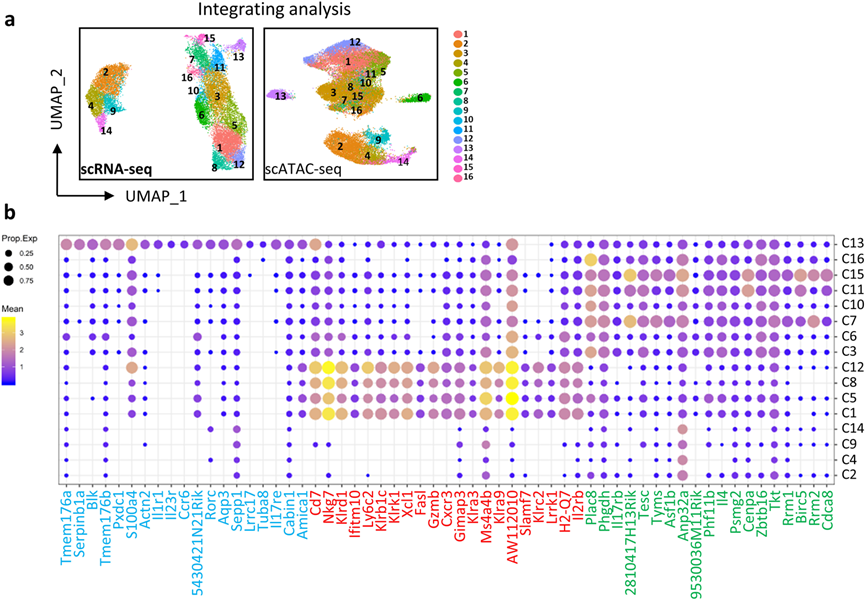
圖2 不同的iNKT簇被分配到功能子集中
2、ST0中的兩個發育軌跡
胸腺中iNKT細胞的發育依賴于位于胸腺皮質的大約1000個ST0 iNKT前體細胞(iNKTp)。在這里,作者鑒定了ST0中的四個簇(C2、C4、C9和C14)(圖1d和3a),并突出顯示了每個簇的特定特征(圖3b)。根據CD4和CD8的表達,這些ST0簇可以分為三組:C14是CD4+CD8+(DP)iNKTp;C9是CD4+CD8?(CD4SP)iNKTp,富含T淋巴細胞存活調節因子,包括Id2和Il7r;C2和C4是CD4?CD8?(DN)iNKTp,其中C4高度表達Cd5和Cd6,而C2 iNKTp豐富表達Egr2和Slamf6,它們分別通過調節PLZF表達和TCR信號強度,對iNKT細胞的發育至關重要(圖3b–d)。盡管先前的研究聲稱iNKT細胞要么是CD4?CD8?(DN),要么是CD4+CD8?(CD4SP),但在ST0中確實發現了一個小的CD4+CD8+(DP)iNKTp簇(C14)(圖3c–e)。
將ST0 iNKTp組織成一個偽時間軌跡。鑒別了兩個潛在的發育分支,即C14-C2-C4分支(稱為DP-DN分支)和C14-C9分支(DP-CD4SP分支)(圖3f)。這兩個分支最終在表達趨化因子受體Ccr7增加的發育末端相遇(圖3g),Ccr7在ST0 iNKTp從胸腺皮質遷移到髓質中是必需的。通過流式細胞儀在蛋白質水平上進一步確認了DP、DN和CD4SP iNKTp中Ccr7的表達模式(圖3h)。因此, DP iNKTp下調CD8或同時下調CD4和CD8的表達,以啟動在胸腺皮質中的DP-CD4SP或DP-DN發育程序,最終遷移到胸腺髓質。
3、ST0中功性能iNKT亞型譜系
接下來,想知道ST0中這兩個發育程序如何對iNKT亞型譜系產生影響。盡管PLZF被注釋為關鍵的iNKT2標志,但它對整體iNKT細胞的發育也至關重要,包括iNKT1和iNKT17細胞。因此,首先評估了Zbtb16(編碼PLZF)在ST0 iNKTp中的表達模式和染色質可及性。發現在DP iNKTp(C14)中,Zbtb16的+40?kb區域和TSS是不可及的,但在CD4SP(C9)和DN(C2和C4) iNKTp中是可及的,其中C2的開放性水平遠高于C4,這與Zbtb16的表達模式非常相似(圖3i,j)。偽時間軌跡和流式細胞儀進一步表明,Zbtb16在DN和CD4SP iNKTp中富集(圖3j,k)。鑒于iNKT2分化需要比iNKT1和iNKT17更強的TCR信號,因此CD4SP中的PLZF高表達iNKTp(C9)更有可能進入iNKT2細胞譜系。
RORγt是調控iNKT17分化的關鍵轉錄因子,但也高度表達于未被信號刺激的DP胸腺細胞,并促進iNKT細胞選擇。與Zbtb16不同,在+4?kb區域的Rorc(編碼RORγt)在DP(C14)和DN(C2和C4)iNKTp中是可及的,但在CD4SP(C9)簇中不可及,而+13?kb到Rorc只在DP iNKTp(C14)中可及。Rorc的表達模式與每個簇中的染色質可及性狀態一致(圖3l)。Rorc在DP iNKTp中顯著高表達,并在DN分支(C4和C2)中逐漸下調,但在CD4SP(C4)中幾乎不可檢測(圖3m),這在流式細胞儀中得到了進一步驗證(圖3n)。總體而言,基于偽時間的發育軌跡分析揭示了ST0可能存在兩個潛在的發育程序,在這兩個程序中,iNKT細胞可能啟動對iNKT2和iNKT17細胞的發育,這早在ST0發生,而iNKT1細胞可能在ST0后啟動。然而,這一假設仍在進一步調查中。
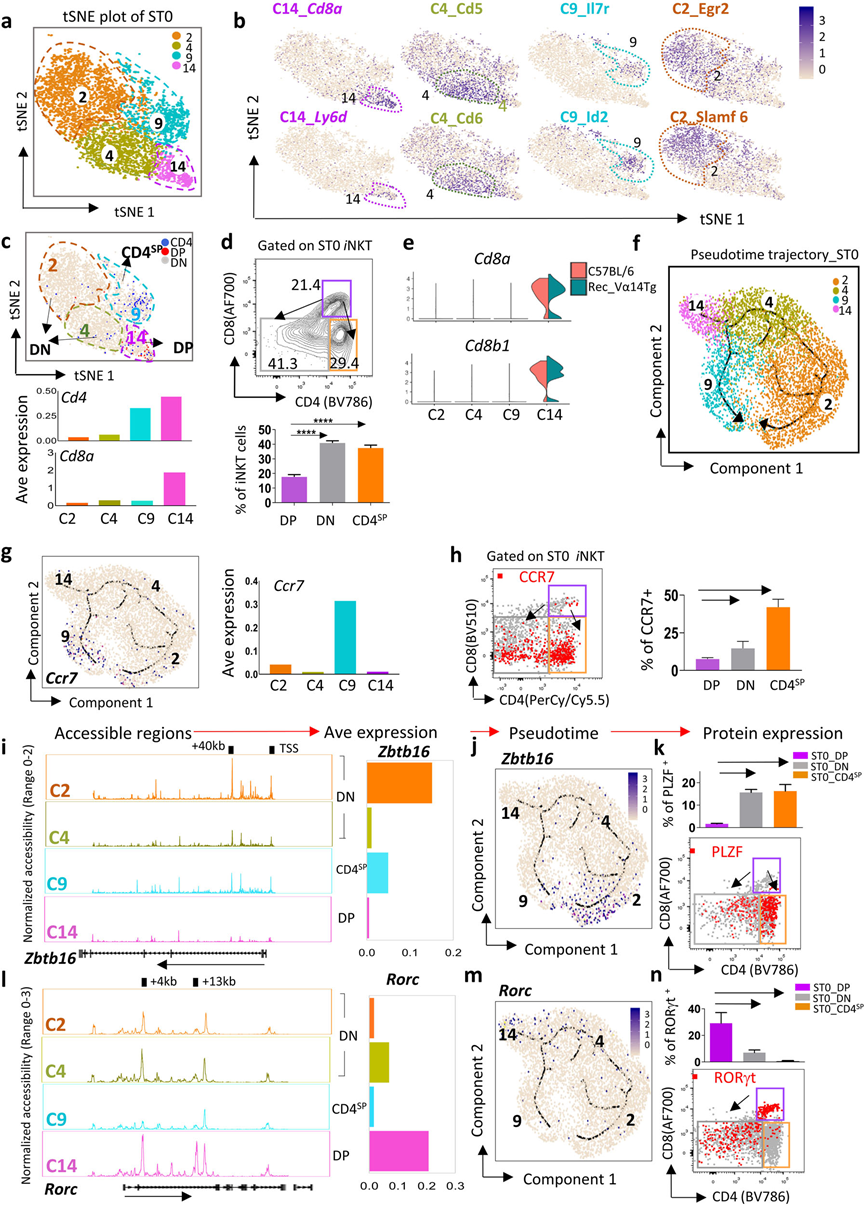
圖3 ST0時期iNKT細胞的細胞多樣性
4、iNKT2細胞的細胞多樣性
為了解ST0后iNKT子集的發展,將聚集的胸腺iNKT細胞(ST0 - 3)映射到偽時間軌跡中(圖4a)。iNKT2細胞展示了廣泛的多樣性,包括C3、C6、C7、C10、C11、C15和C16簇(圖2b和4a, b)。首先評估了整合的iNKT細胞簇(C1–C16)中Zbtb16染色質的可及性。在Zbtb16位點的181 kb內,發現iNKT2、iNKT1和iNKT17簇中高度可訪問的區域,但在ST0簇(C2、C4、C9和C14)中區域較弱。Zbtb16的表達與其染色質可及性密切匹配(圖4c)。iNKT2簇和ST0中的C2和C9顯示了GATA3結合活性的高活性,GATA3對iNKT2分化至關重要(圖4d)。正如預期的那樣,對于iNKT2細胞的大多數標志基因,它們在中間階段逐漸增加,然后在終端分化期間下調。然而,一些基因,包括Btg1、Cebpb和Osgin1,僅在iNKT發育末端附近出現,這可能與iNKT2細胞的終末事件相關(圖4e)。
先前的研究表明,ST1和ST2中的iNKT細胞經歷高度的增殖。細胞周期通路富集分析表明,iNKT2簇(C7、C11和C15)表現出高度增殖的特征,這些特征可能處于S期(C7)或G2/M期(C11和C15)(圖4f)。Ingenuity Pathway Analysis(IPA)表明,簇C3、C6、C10和C16在功能上存在差異。C16是從ST0到ST1的iNKT細胞的短暫階段,這些細胞迅速上調了與糖酵解和氧化磷酸化都相關的基因的表達(圖4g),表明它們對能量的需求增加。C6在ST1中終止,這些細胞富集了與TCR信號、共刺激信號、細胞骨架、TNFR以及細胞死亡和衰竭通路相關的基因(圖4g)。總體而言,iNKT2展現出極大的細胞多樣性。
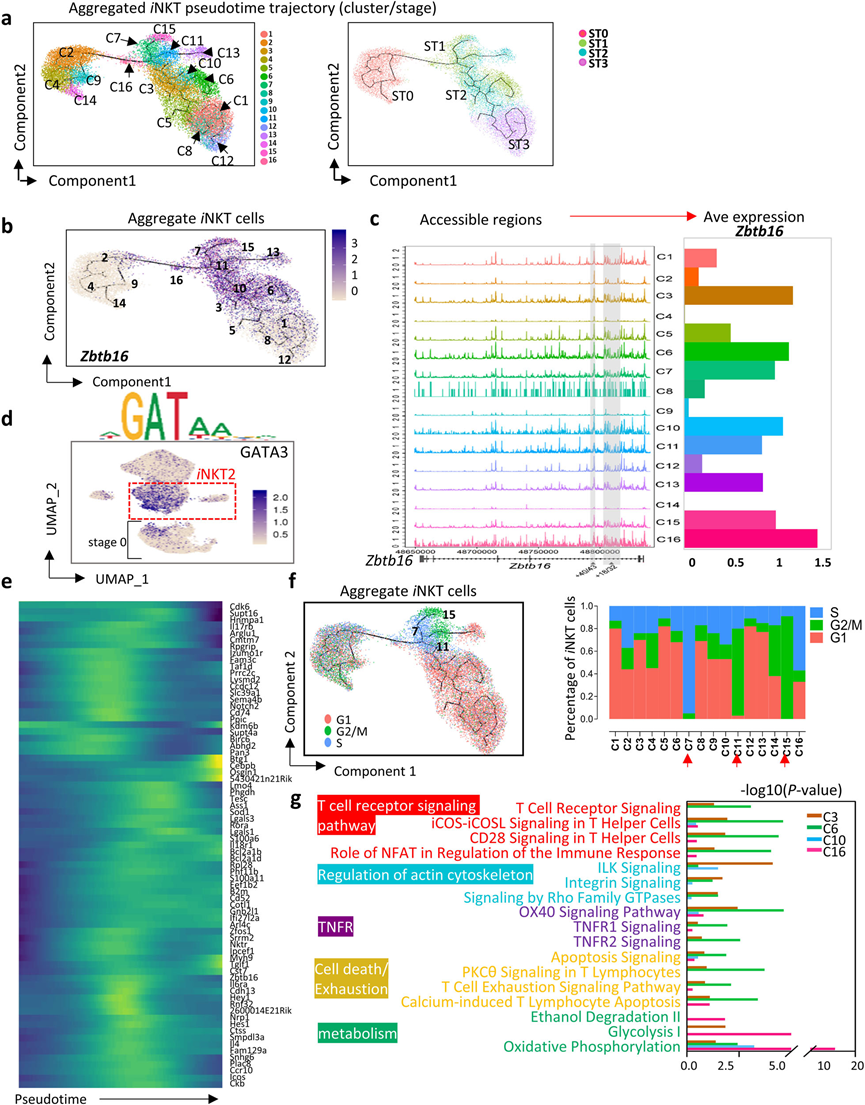
圖4 iNKT2細胞的細胞多樣性
5、iNKT1細胞的異質性
iNKT1細胞分化受轉錄因子Tbx21的調控,分為四個簇(C1、C5、C8和C12)(圖2b和5a)。在Tbx21位點的17.2 kb內,啟動子區域–3 kb和–4 kb在C1、C5和C12中高度可訪問,但在C8中較不可訪問(圖5b)。這些相同的區域在ST1和ST2中的高增殖C7、C11和C15 iNKT2簇中也是可訪問的。重要的是,Tbx21結合位點的活性也發生在這些簇中,而在ST0簇中則不是如此(圖5c)。流式細胞儀分析進一步證實,ST1和ST2中的PLZFhiiNKT2細胞中有一小部分表達T-bet,并且通過Ki-67測量具有相當的增殖能力(圖5d)。因此,iNKT1的前體可能隱藏在這些所謂的高增殖PLZFhiT-bethi iNKT2細胞中。
偽時間軌跡分析發現一個新的標志基因,Slamf7,在iNKT1細胞簇中富集(圖5e)。流式細胞儀進一步證實了這一點,基于PLZFloT-bethi iNKT1細胞中SLAM7和NK1.1表達之間的強相關性(圖5f)。與其他iNKT1細胞簇相比,NK細胞相關的標志基因Slamf4進一步區分了末端的C12。SLAMF4+ iNKT細胞(C12)主要是DN,逐漸從PLZFloT-bethi iNKT1細胞中分化出來(圖5g–i)。SLAMF4+ iNKT1細胞主要分泌IFN-γ,IL-4較少,類似于“經典iNKT1細胞”,而大多數SLAMF4? iNKT1細胞在刺激后分泌IL-4和IFN-γ(圖5j)。C5細胞富集了參與干擾素信號通路的Ifit1和Ifit3(圖5k)。總體而言,iNKT1細胞從ST1開始,起初表現為目前定義的“iNKT2細胞”,逐漸完成分化并最終轉化為末端的SLAMF4+ iNKT1細胞,即經典的分泌IFN-γ的iNKT1細胞。
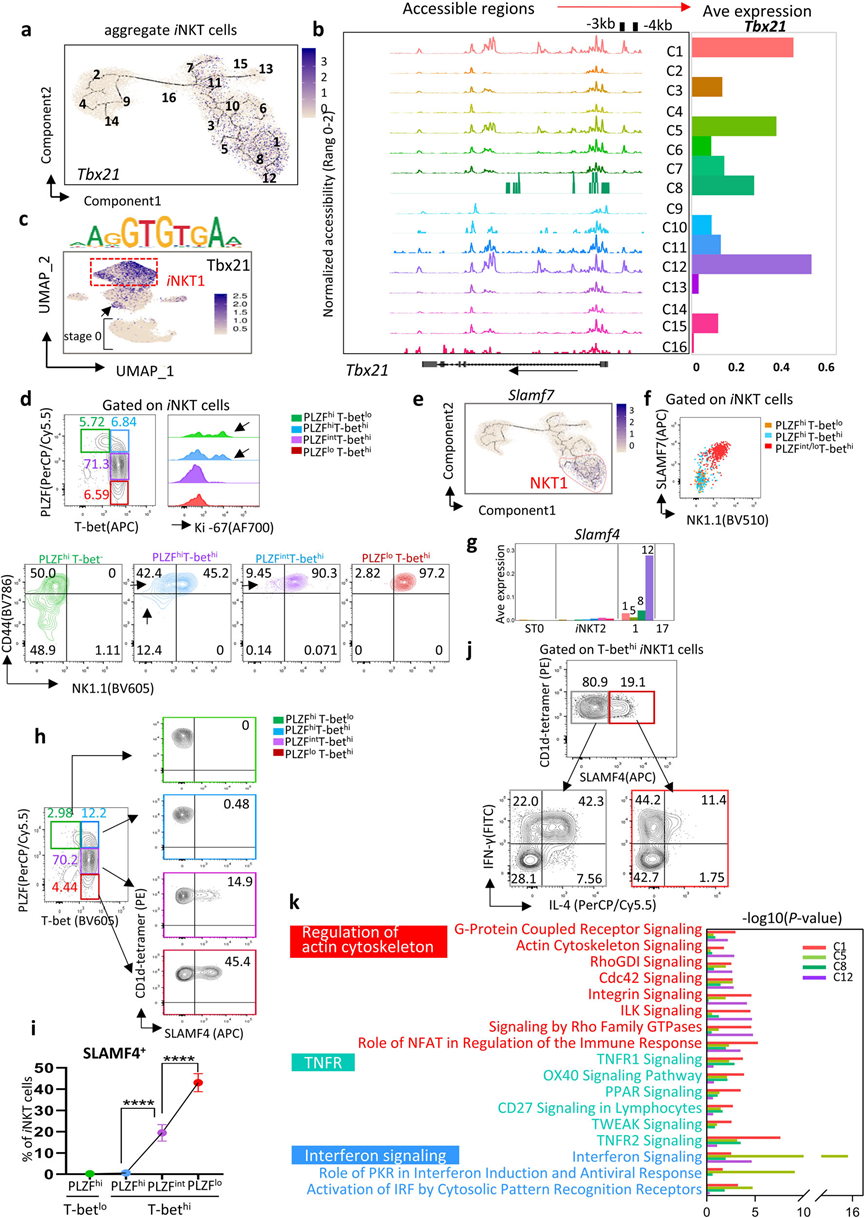
圖5 iNKT1細胞具有廣泛的細胞異質性
6、iNKT17細胞表現出有限的多樣性
C13被指定為iNKT17細胞,它在圖2b和6a中與其他簇明顯分開。觀察到Rorc的+4?kb和+14?kb區域在ST0簇(C2、C4和C14)和成熟的iNKT17簇(C13)中是可訪問的(圖6b),并且RORC的結合位點在iNKT17簇以及ST0 iNKTp中也被激活(圖6c)。這些數據表明,iNKT細胞可能在ST0開始其iNKT17分化,并在C13完成其分化。在mRNA水平上,Rorc在ST0的C14中高度表達,與Rorc的染色質可及性一致,但在C4和C2中逐漸下調,并在C13中重新上調,表明其他轉錄因子可能靶向開放的Rocr位點,并在iNKT17分化過程中調控Rorc的表達。
為追蹤iNKT17細胞的發育軌跡,進一步在一個有序的iNKT細胞軌跡中檢查了iNKT17標志基因,并發現少數iNKT17標志基因,包括Rorc,在早期iNKTp中最初表達,然后在它們達到成熟的iNKT17之前。然而,大多數與iNKT17相關的標志基因在iNKTp中幾乎不表達(圖6d)。進一步發現一個新的標志基因, Aqp3,在胸腺iNKT17(PLZFintRORγt+)細胞中特異表達(圖6e,f)。總體而言,iNKT細胞可能在ST0啟動iNKT17分化,而這些iNKT17細胞表現出有限的多樣性。
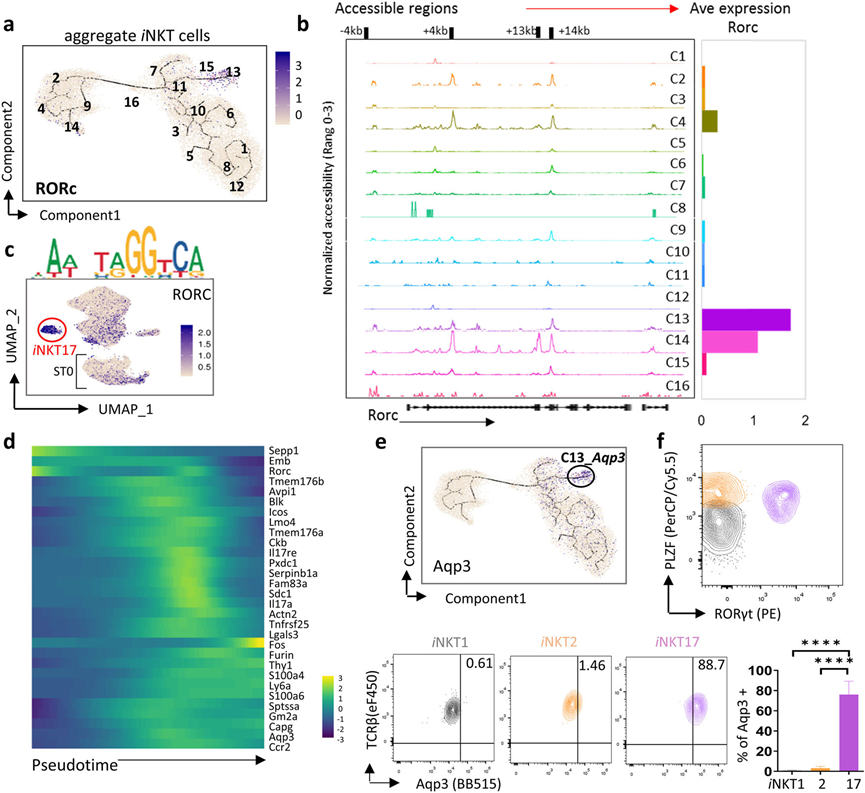
圖6 iNKT17細胞表現出有限的異質性
7、Cbfβ調控iNKT細胞的早期分化
先前的研究指出,Egr2和Slamf6通過調節Zbtb16的表達和TCR在ST0中控制早期iNKT細胞的發育。在這里,作者發現了一個新的共轉錄因子,Cbfβ,它與Egr2和Slamf6表現出相似的表達模式,并在iNKT細胞的ST0階段表現出很高的富集,特別是在DP-DN分支C2中(圖7a, b)。
Cbfβ編碼的CBFB是一個非DNA結合的調節亞基,通過異構增強RUNX(RUNX1、RUNX2和RUNX3)的特異性DNA結合能力,從而調節它們目標基因的轉錄。為確定Cbfβ是否調節iNKT發育,檢查了胸腺特異性Cbfβ敲除小鼠(CD4CreCbfβ f/f,Cbfβ KO)中的iNKT細胞發育,其中Cbfβ1和Cbfβ2均被敲除。Cbfβ敲除導致胸腺iNKT細胞的頻率和絕對數量顯著減少(圖7c)。進一步更全面的分析揭示了Cbfβ KO小鼠中ST2和ST3 iNKT細胞的選擇性和顯著減少。Cbfβ KO小鼠中ST0和ST1 iNKT細胞的頻率顯著增加,但在Cbfβ KO和WT對照組之間,絕對數量是可比的(圖7d)。有趣的是,在Cbfβ KO小鼠的ST0 iNKT細胞中,DP iNKTp增加(圖7e),表明Cbfβ的敲除部分阻斷了DP iNKTp向CD4SP或DN譜系的轉化。作者推測Cbfβ可能影響DP階段的iNKT細胞選擇。
ST1 iNKT細胞經歷迅速的增殖,并包含具有高Zbtb16編碼PLZF表達的iNKT亞群的前體。在這里,觀察到在Cbfβ KO iNKT細胞中,ST1時的PLZF表達顯著下調(圖7f)。在一個增殖標記Ki-67上也觀察到了類似的現象(圖7g)。這些數據表明,Cbfβ缺乏的iNKT細胞進入了相對靜止狀態,無法在ST1正常上調PLZF表達。此外,在ST1中殘留的PLZF+ iNKT細胞未能共同表達T-bet以啟動iNKT1分化,也未能共同表達RORγt以啟動iNKT17分化(圖7h,i)。總體而言,研究表明,Cbfβ在控制ST0時的早期iNKT細胞發育、ST1/2時的iNKT細胞分化以及ST3時的最終成熟中起到關鍵調節作用(圖7j)。
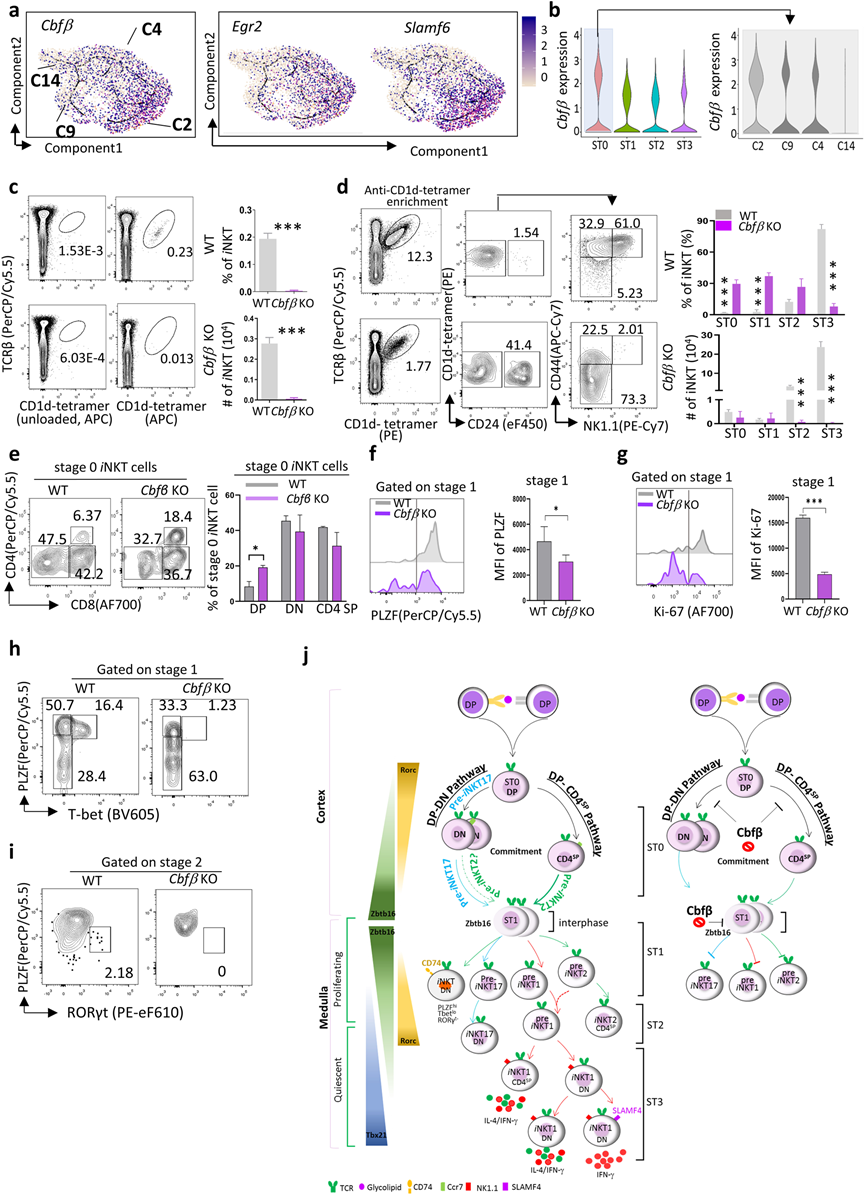
圖7 Cbfβ調控iNKT細胞早期細胞譜系
實驗方法:
構建條件性敲除小鼠,流式細胞術,骨髓嵌合體移植實驗,小鼠iNKT細胞富集和分選,qRT-PCR,WB,scRNA-seq,iNKT scRNA-seq數據集與參考文獻的比較分析,scATAC-seq,scRNA-seq和scATAC-Seq的整合分析
參考文獻:
Wang J, Adrianto I, Subedi K, Liu T, Wu X, Yi Q, Loveless I, Yin C, Datta I, Sant'Angelo DB, Kronenberg M, Zhou L, Mi QS. Integrative scATAC-seq and scRNA-seq analyses map thymic iNKT cell development and identify Cbfβ for its commitment. Cell Discov. 2023 Jun 20;9(1):61. doi: 10.1038/s41421-023-00547-x. PMID: 37336875; PMCID: PMC10279728.