






T淋巴細胞誘導癌細胞分泌半乳凝素-9,與其他免疫檢查點蛋白合作,促進免疫抑制
半乳糖凝集素-9是凝集素蛋白家族的一員,對人體免疫反應起著至關重要的調節作用,特別是因為它具有抑制T淋巴細胞和自然殺傷細胞抗癌活性的能力。最近的證據表明,半乳糖凝集素-9在廣泛的人類惡性腫瘤中高表達,包括最具侵襲性的腫瘤,如高級別膠質母細胞瘤和胰腺導管腺癌,以及常見的惡性腫瘤,如乳腺癌、肺癌和結直腸癌。然而,實體腫瘤細胞要么分泌極少量的半凝集素-9,要么在大多數情況下根本不分泌它。作者的目的是闡明T細胞是否可以誘導來自實體惡性腫瘤的人類癌細胞分泌半乳糖凝集素-9,以及與細胞表面蛋白相比,這種可溶性形式是否表現出更高的全身免疫抑制活性。作者的結果強調了半乳糖凝集素-9在抗癌免疫逃避中的關鍵作用。因此,半乳糖凝集素-9和控制其產生的調控途徑應被視為大量癌癥免疫治療的關鍵靶點。該文發布于2023年1月《Journal of ImmunoTherapy of Cancer》,IF=10.9。
技術路線:
主要研究結果:
1、T淋巴細胞激活源自實體惡性腫瘤的人癌細胞的半乳糖凝集素-9分泌
作者研究了T細胞是否能誘導來自實體惡性腫瘤的人癌細胞分泌半乳糖凝集素-9。作者研究了那些自身不分泌半乳糖凝集素-9的細胞和那些分泌少量這種蛋白質的細胞。此外,作者還測試了非惡性快速增殖的人類細胞,包括HaCaT角質形成細胞和原代人類胚胎細胞。所有被測試的細胞都清晰地表達了可檢測到的且通常含量很高的半凝集素-9。腫瘤細胞模型選用MCF-7乳腺癌細胞、WT3ab Wilms腫瘤細胞、G401腎橫紋肌樣腫瘤細胞和LN-18高級別膠質母細胞瘤細胞。所有細胞與Jurkat T細胞按1:1的比例共培養16小時。然后分離Jurkat T細胞,用新鮮培養基洗滌2-3次,然后用各自的細胞特異性培養基孵育共培養的細胞。細胞培養4小時,分別于2、3、4小時檢測半凝集素-9的分泌。分離后,Jurkat T細胞分泌的半凝集素-9幾乎無法檢測到。人類癌細胞分泌高水平的半乳糖凝集素-9,HaCaT角質形成細胞分泌較少,人類胎兒細胞產生幾乎無法檢測到的蛋白質(圖1A)。作者早期的研究證實了半乳糖凝集素-9在MCF-7、WT3ab、HaCaT和原代人胚胎細胞中的表達。然后作者研究了不同類型的T細胞是否能誘導半乳糖凝集素-9的分泌。為了進行比較,作者將MCF-7人乳腺癌細胞與Jurkat T細胞(cd4陽性)、TALL-104細胞毒性(cd8陽性)淋巴細胞以及原代人cd3陽性T細胞共培養。所有共培養按1:1的比例進行16小時,然后按上述方法處理。分離后,所研究的T細胞均未分泌半乳糖凝集素-9 (Jurkat T細胞除外,其分泌量幾乎無法檢測到(<100 pg/mL))。然而,在所有三種情況下,MCF-7細胞都分泌大量的半凝集素-9(圖1B)。如上所述,在所研究的貼壁細胞中,在沒有T淋巴細胞的情況下,只有LN-18能夠在靜止狀態下分泌可檢測到(但含量很低)的半凝集素-9(圖1C)。然后作者研究了CD4或CD8蛋白是否參與T細胞誘導的半凝集素-9分泌。作者將原代人T細胞與MCF-7細胞在CD4或CD8中和抗體存在或不存在的情況下共培養16小時。然后,如上所述,分離T細胞,然后在條件培養基中測量培養的MCF-7細胞在2、3和4小時后釋放的半凝集素-9水平。在MCF-7和原代人T細胞共培養16小時的培養基中也測量了半乳糖凝集素-9。細胞上Western分析證實了抗CD4和抗CD8與原代T細胞的結合。作者發現這兩種抗體都不能顯著減少t細胞誘導的MCF-7細胞中半乳糖凝集素-9的分泌,這表明CD4和CD8不太可能參與這一過程。然而,由于作者看到抗CD8抗體引起的效果略有降低,作者將高活性的LN-18膠質母細胞瘤細胞與Jurkat T(僅表達CD4而不表達CD8蛋白)細胞共培養16小時,如上所述。然后分離Jurkat T細胞,LN-18細胞培養4小時,2小時、3小時和4小時后在條件培養基中測量半凝集素-9水平。在LN-18和Jurkat T細胞共培養16小時的培養基中也測量了半乳糖凝集素-9。作者證實抗cd4在Jurkat T細胞上高表達,而使用細胞上的Western分析幾乎檢測不到抗cd8。此外,作者用流式細胞術測量了共培養后Jurkat T細胞中的CD4和CD8,發現它們CD4陽性,而CD8水平幾乎無法檢測到。CD4和CD8中和抗體對T細胞誘導的LN-18細胞半凝集素-9分泌無影響。在抗cd8抗體存在的情況下,觀察到半乳糖凝集素-9分泌非常輕微(不顯著)的減少,與MCF-7細胞相似。然而,由于Jurkat T細胞中缺乏CD8表達,這不太可能是特異性的。重要的是,FACS分析證實共培養后LN18細胞中沒有任何CD4的痕跡。MCF-7細胞只表達一種半乳糖凝集素-9結合受體- Tim-3,16 16 WT3ab細胞只表達T細胞活化抑制因子(VISTA)蛋白和微量的Tim-3,16而LN-18細胞同時表達Tim-3和VISTA。作者評估了半乳糖凝集素-9是否在復合物中與其受體片段一起釋放,這些片段可以作為該蛋白的載體/轉運者。作者免疫沉淀半乳糖凝集素-9,在不煮沸樣品的情況下使用Western blot分析檢測。當MCF-7細胞與Jurkat T細胞共培養時,這導致半乳糖凝集素-9以Tim-3片段復合物的形式分泌(圖2A)。然而,在WT3ab細胞中,它以與VISTA片段復合物的形式分泌(圖2B)。與Tim-3不同,VISTA在SDS-PAGE電泳中與半乳糖凝集素-9分離,證實了作者之前報道的Tim-3與VISTA相比對半乳糖凝集素-9具有更高親和力的發現。這兩種受體都存在于LN-18中,其中Tim-3是半乳糖凝集素-9的載體(圖2C),這與作者之前對同樣表達Tim-3和VISTA受體的原代人胚胎細胞的發現一致,作者希望評估MCF-7和細胞毒性TALL-104細胞之間的相互作用,以發現MCF-7衍生的半凝集素-9是否能夠調理細胞毒性T細胞。MCF-7和TALL-104細胞按1:1的比例共培養,然后按上述方法分離。裂解TALL-104細胞,從靜止的TALL-104裂解物中裝載約50-60μg/孔的細胞總蛋白,用于Western blot檢測半乳糖凝集素-9(這些細胞表達低量的半乳糖凝集素-9和Tim-3)。然后,作者在與T細胞共培養后裂解的TALL-104細胞中加載較少的蛋白(約15μg/孔)。結果,在與MCF-7細胞共培養的TALL104細胞的裂解物中,作者觀察到抗tim -3和抗半乳糖凝集素-9抗體均可檢測到約52 kDa的條帶(圖3A)。這與MCF-7細胞分泌的半乳糖凝集素-9相對應(圖3B),在半乳糖凝集素-9免疫沉淀后的培養基中檢測到。重要的是,該復合物對TALL-104細胞表面受體的親和力低于對運輸它的Tim-3的親和力。作者可以排除這不是TALL104細胞產生的內部復合物,因為已知該內部復合物在Tim-3脫落前的分子量約為70 kDa。另外,作者發現TALL-104與MCF-7細胞共培養后,半凝集素-9 mRNA水平顯著下調。VISTA mRNA水平也有相同的觀察結果,而Tim-3 mRNA水平顯著上調(詳見圖3C)。在MCF-7細胞中,Tim-3和半乳糖凝集素-9 mRNA水平均顯著上調(圖3D)。同時,TALL-104細胞相對不能將顆粒酶B傳遞到MCF-7細胞中(圖3E, F),這與作者之前的觀察結果一致。
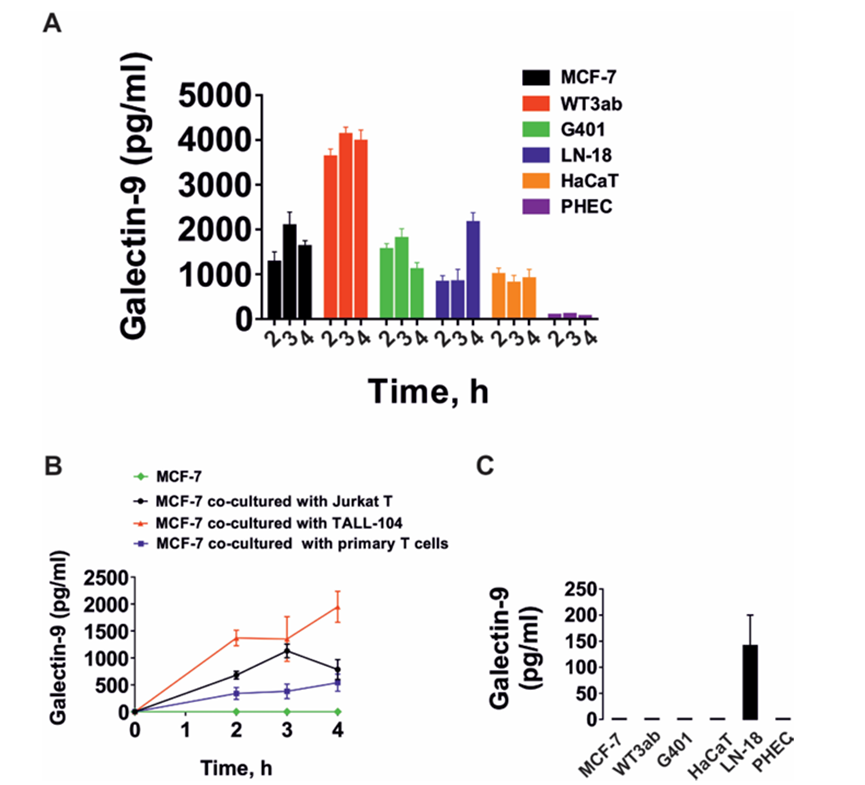
圖1、T細胞誘導來自實體惡性腫瘤和角質形成細胞的人癌細胞分泌半凝集素-9
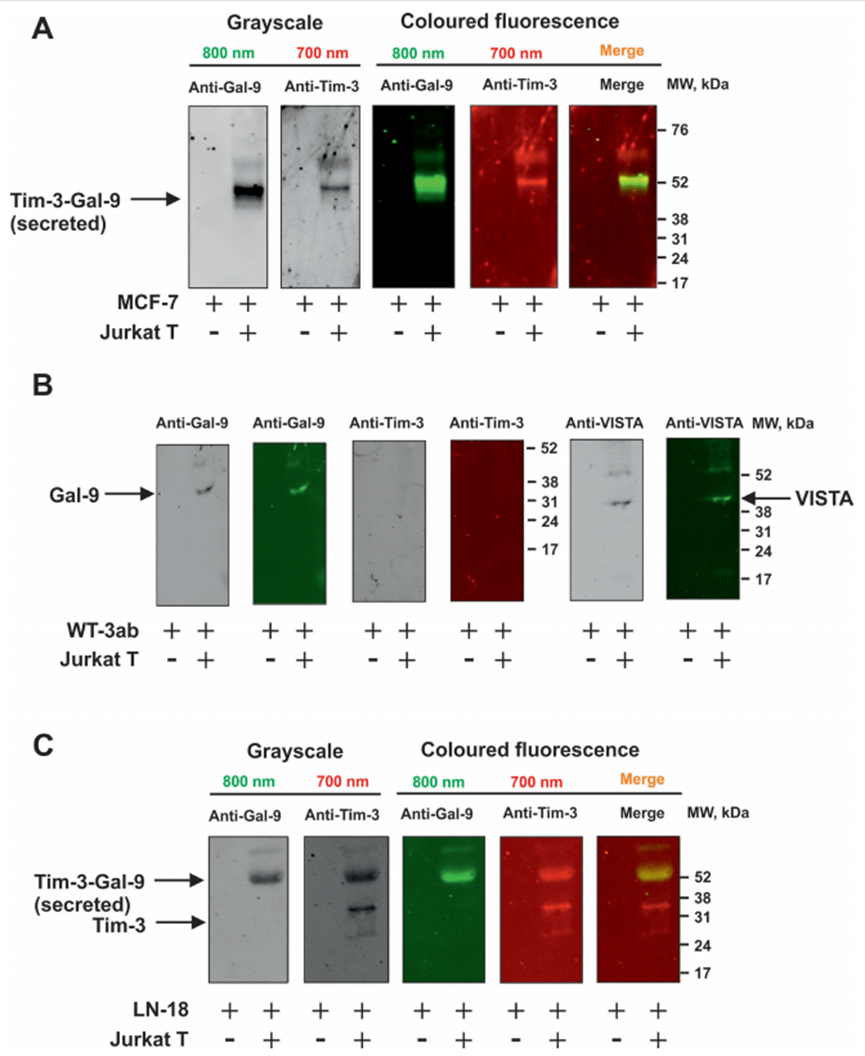
圖2、Tim-3或VISTA在分泌過程中干擾半乳糖凝集素-9
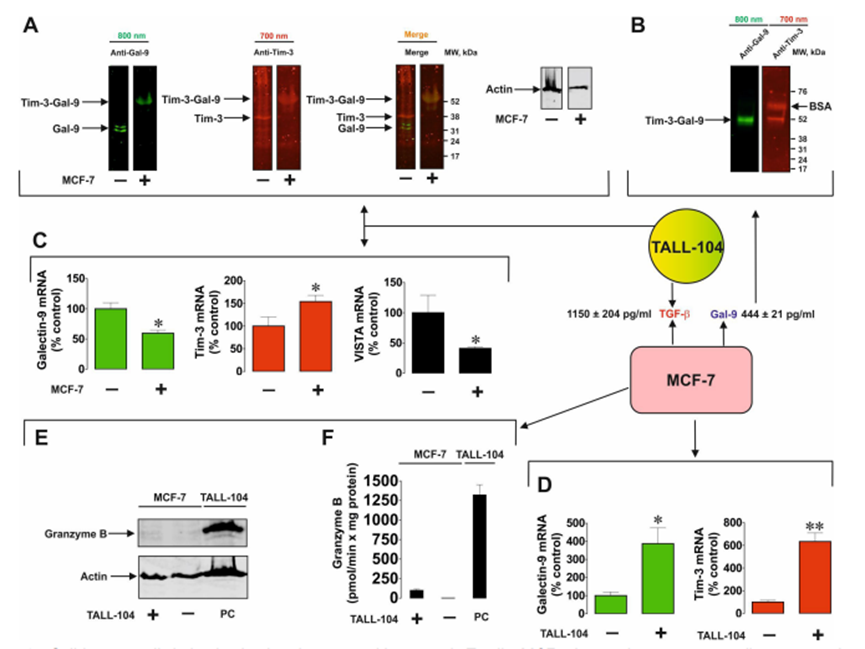
圖3、實體腫瘤細胞來源的半乳糖凝集素-9與細胞毒性T細胞相互作用。MCF-7人乳腺癌細胞與TALL-104按1:1的比例共培養16小時
2、來源于實體惡性腫瘤的人癌細胞在T細胞誘導下有兩種半乳糖凝集素-9分泌機制
作者考慮了來自實體惡性腫瘤的人類癌細胞T細胞誘導半乳糖凝集素-9分泌的兩種不同機制的可能性。作者將MCF-7和Jurkat T細胞按1:1的比例共培養16小時,然后如上所述分離它們,隨后將MCF-7細胞在新鮮培養基中培養4小時,在2、3和4小時時測量半凝集素-9的分泌。細胞在不同抑制劑存在或不存在的情況下共培養。其中包括U73122 (PLC抑制劑)、G?6983 (PKC抑制劑)、液泡素-1(溶酶體胞吐抑制劑)、AZD2014 (mTOR抑制劑)、EDTA(基質金屬蛋白酶抑制劑)和含有溶酶體蛋白酶和基質金屬蛋白酶阻滯劑的蛋白酶抑制劑混合物(PIC)(圖4A)。作者發現G?6983、液泡素-1、EDTA和PIC下調分泌(圖4B)。AZD2014上調了這種分泌,這表明它是自噬依賴性溶酶體分泌(液泡素-1的抑制作用證實了這一點),而不像AML細胞中半乳糖凝集素-9的分泌是自噬依賴性的。這些結果表明半乳糖凝集素-9的分泌有兩種機制。第一個是該蛋白易位到細胞表面,然后通過基質金屬蛋白酶(如AML細胞)從細胞表面脫落。另一種是自噬體的形成,隨后是溶酶體胞吐作用,其中半乳糖凝集素-9也脫落。這一結論是基于PIC阻斷溶酶體蛋白酶和基質金屬蛋白酶后半乳糖凝集素-9的分泌減少。參與該過程的PKC最有可能是鈣依賴性和二酰基甘油(DAG)非依賴性的(PLC對分泌過程沒有影響)。作者通過將MCF-7與Jurkat T細胞以1:1的比例共培養2小時,然后使用Fluo4活性染料檢測細胞內鈣水平,檢查MCF-7細胞內鈣水平是否上調,方法見材料和方法部分。作者發現,與靜止的MCF-7細胞相比,與Jurkat T細胞共培養的MCF-7細胞的細胞內鈣水平顯著上調。這表明環境支持激活Ca2+依賴性PKC異構體,該異構體與該酶的DAG/Ca2+依賴性異構體G?6983一起被抑制。
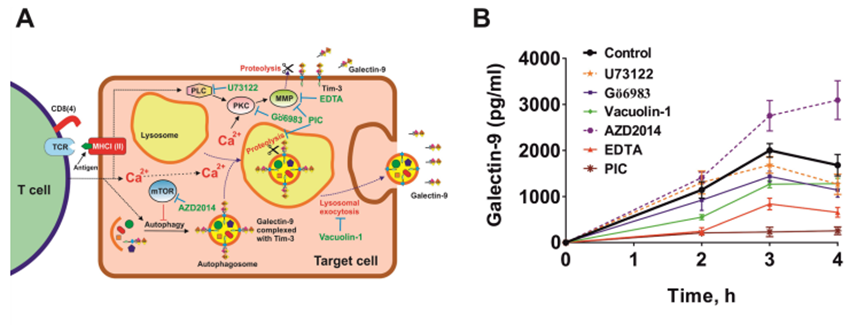
圖4、T細胞誘導MCF-7人乳腺癌細胞分泌半乳糖凝集素-9可通過兩種機制發生。
3、半凝集素-9可以協同VISTA抑制T淋巴細胞的抗癌活性
然后作者研究了半乳糖凝集素-9的免疫抑制活性,以及它如何與其他免疫檢查點合作以實現T細胞失活。首先,作者將MCF-7人乳腺癌細胞與Jurkat T細胞(具有T輔助活性的CD4陽性細胞)以1:1的比例共培養16小時,無論是否存在9S2-1半乳糖凝集素-9中和(阻斷)抗體或同型對照抗體。在孵育完成后,作者在細胞培養基和Jurkat T細胞裂解物中測量了IL-2。作者還分析了Jurkat T細胞裂解物中磷脂酰肌醇3-激酶(PI-3K)的活性(PI-3K活性及其途徑有助于輔助性T細胞中IL-2的表達,并表明TCR信號的激活)。重要的是,靜止的Jurkat T細胞分泌的IL-2幾乎無法檢測到,而MCF-7則不分泌任何IL-2。作者發現,Jurkat T細胞在與MCF-7細胞相互作用過程中產生的分泌(但不是細胞相關IL-2的水平)以及IL-2的總量(分泌+在裂解物中檢測到的細胞相關水平)在半乳糖凝集素-9中和抗體而不是同型對照抗體的存在下顯著降低(圖5A)。對PI-3K也觀察到同樣的效果。這意味著半乳糖凝集素-9支持PI3K在Jurkat T細胞中的激活,并通過它們增強IL-2的產生。然后,作者將Jurkat T細胞與LN-18(與MCF-7細胞不同,LN-18也表達VISTA)以1:1的比例共培養16小時。作者發現,在LN-18的存在下,Jurkat T細胞并不比靜止的Jurkat T細胞分泌更多的IL-2。然而,轉化生長因子-β 1型(TGF-β)和半乳糖凝集素-9的分泌水平高度上調(圖5B)。與作者之前的研究結果一致,作者發現TGF-β在mRNA和蛋白水平上上調了LN-18細胞中半乳糖凝集素-9和VISTA的表達。然后,作者將LN-18細胞與原代人cd -3陽性T細胞共培養,并評估IL-2、顆粒酶B、TGF-β和半乳糖凝集素-9在培養基中的釋放。作者發現顆粒酶B和IL-2的釋放水平沒有增加,但TGF-β的產生和半乳糖凝集素-9的分泌均顯著上調(圖5C)。重要的是,當LN-18細胞與Jurkat T細胞或原代人T淋巴細胞共培養時,細胞表面的VISTA水平顯著上調。ELISA未檢測到LN-18分泌的VISTA。為了評估VISTA對半乳糖凝集素-9在T細胞中觸發的事件的貢獻,作者在缺乏或存在半乳糖凝集素-9或vistan中和抗體的情況下共培養LN-18和Jurkat T細胞。作者發現,當半乳糖凝集素-9被中和時,IL-2的分泌(與MCF-7細胞存在時不同)沒有改變,但當VISTA被中和時,IL-2的分泌會上調(圖6A)。在Jurkat T細胞裂解液中也觀察到同樣的效果。這意味著VISTA有助于抑制T輔助活性。為了確認PI-3K在IL-2產生中的作用,作者在共培養中加入了30μM LY294002 (PI-3K抑制劑),發現PI-3K的活性和IL-2的釋放都減弱了(圖6A)。重要的是,經FACS分析證實,Tim-3存在于與Jurkat T細胞共培養的LN-18細胞表面,并且與半乳糖凝集素-9的表達水平幾乎相同。為了排除Tim-3參與觀察到的效應,作者按照上述方法在存在或不存在Tim-3中和抗體(4BS)的情況下共培養細胞。Tim-3中和對PI-3K活性和IL-2釋放均無影響(圖6A)。下一步是用異種移植模型在體內證實觀察到的效果。首先,為了驗證LN-18細胞是否能夠抑制小鼠T細胞活性,作者在不存在半乳糖凝集素-9或VISTA中和抗體的情況下,以1:1的比例與小鼠T細胞共培養16小時。作者發現共培養導致細胞培養基中釋放的顆粒酶B具有大量活性(圖6B)。當半乳糖凝集素-9中和時,這種活性進一步高度上調,當VISTA中和時,這種活性也顯著上調(圖6B)。當半乳糖凝集素-9或VISTA被中和時,LN-18細胞的活力分別降低(圖6B)。小鼠IL-2分泌只有在VISTA被中和時才會上調,這與LN-18與Jurkat T細胞共培養時的觀察結果一致(圖6A)。小鼠T細胞裂解物中的人半乳糖凝集素-9(總細胞相關半乳糖凝集素-9)通過ELISA(與小鼠半乳糖凝集素-9無交叉反應)進行測量,如材料和方法部分所述。作者發現,當半乳糖凝集素-9被中和時,它在T細胞中幾乎不存在,而當VISTA被中和時,它在小鼠T細胞中的水平顯著下調(圖6B)。VISTA中和的這種效果也可能是半乳糖凝集素-9無法在小鼠T細胞上結合VISTA的結果。
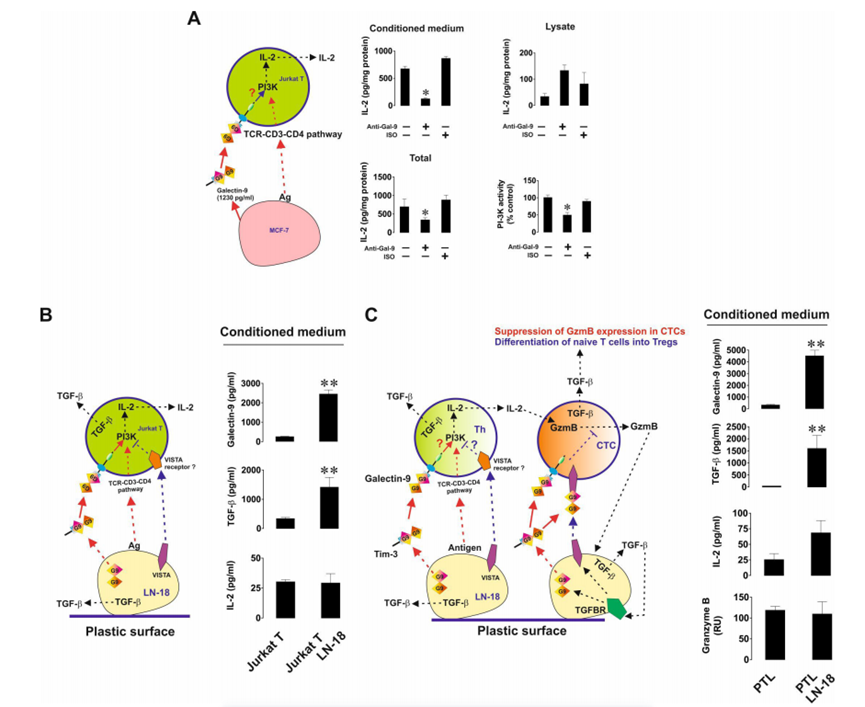
圖5、來源于實體惡性腫瘤的人癌細胞產生的半乳糖凝集素-9觸發輔助性T細胞的PI-3K激活和il - 2產生。表達半乳糖凝集素-9和VISTA的人類癌細胞抑制輔助性和細胞毒性T細胞活性
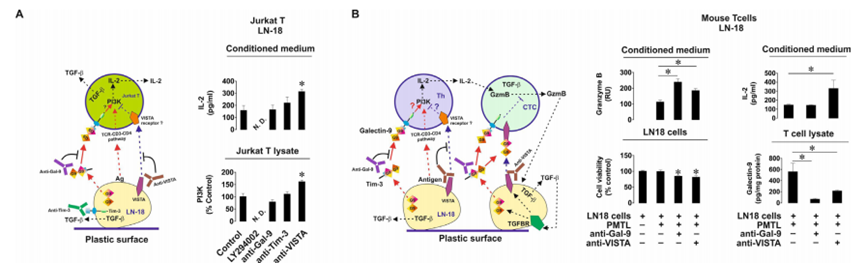
圖6、VISTA與半凝集素-9協同抑制輔助性T細胞和細胞毒性T細胞的活性
4、來源于實體惡性腫瘤的人癌細胞能夠分泌半乳糖凝集素-9,在體內抑制細胞毒性T細胞的活性
為了在體內證實作者的發現,作者使用了C57 BL16小鼠。以5只小鼠作為對照組,其余5只小鼠分別皮下注射2×106 LN-18細胞(具有高度致瘤性),5小時后分析其血液(由于這些小鼠具有免疫活性,并且植入人LN-18具有高度活性,因此在LN-18細胞被小鼠免疫抑制之前,用5小時的時間觀察其最大的免疫逃避反應)。首先,作者分離了LN-18細胞注射區域,并將其置于細胞裂解緩沖液(50 mM Tris pH 7.5, 150 mM NaCl, 5 mM EDTA和0.5% NP-40)中進行軟均質,以溶解分泌到微環境中的蛋白質,并獲得其中存在的裂解的造血細胞。作者發現獲得的勻漿含有高水平的人半乳糖凝集素-9(圖7A),但不含任何人VISTA。在對照組中檢測不到這兩種蛋白。對照組和ln -18注射組。注射微環境(皮下區域,LN-18細胞被注射)含有T細胞,因為在勻漿中可以清楚地檢測到CD3蛋白(圖7A)。注射ln -18的小鼠血漿中含有明顯可檢測到的人半乳糖凝集素-9(對照組未檢測到)。這些小鼠的T細胞裂解物也顯示出高水平的人半乳糖凝集素-9,這在對照動物的T淋巴細胞中檢測不到(圖7B)。此外,血漿TGF-β水平高度上調(圖7B)。作者還評估了人半乳糖凝集素-9是否影響小鼠T淋巴細胞的抗癌細胞毒活性。為此,將小鼠T細胞與100 nM PMA預處理的等量K562細胞以1:1的比例與從對照和注射ln -18的小鼠中分離的小鼠原代T細胞共培養24小時。細胞共孵育24小時,測定K562細胞活力。作者發現,與未與小鼠T淋巴細胞共培養的細胞和與注射了LN-18細胞的小鼠的T細胞共培養的細胞相比,K562細胞與對照組小鼠T細胞共培養的細胞的活力顯著降低(圖7C)。因此,可以得出結論,人半乳糖凝集素-9下調小鼠T淋巴細胞的細胞毒活性。
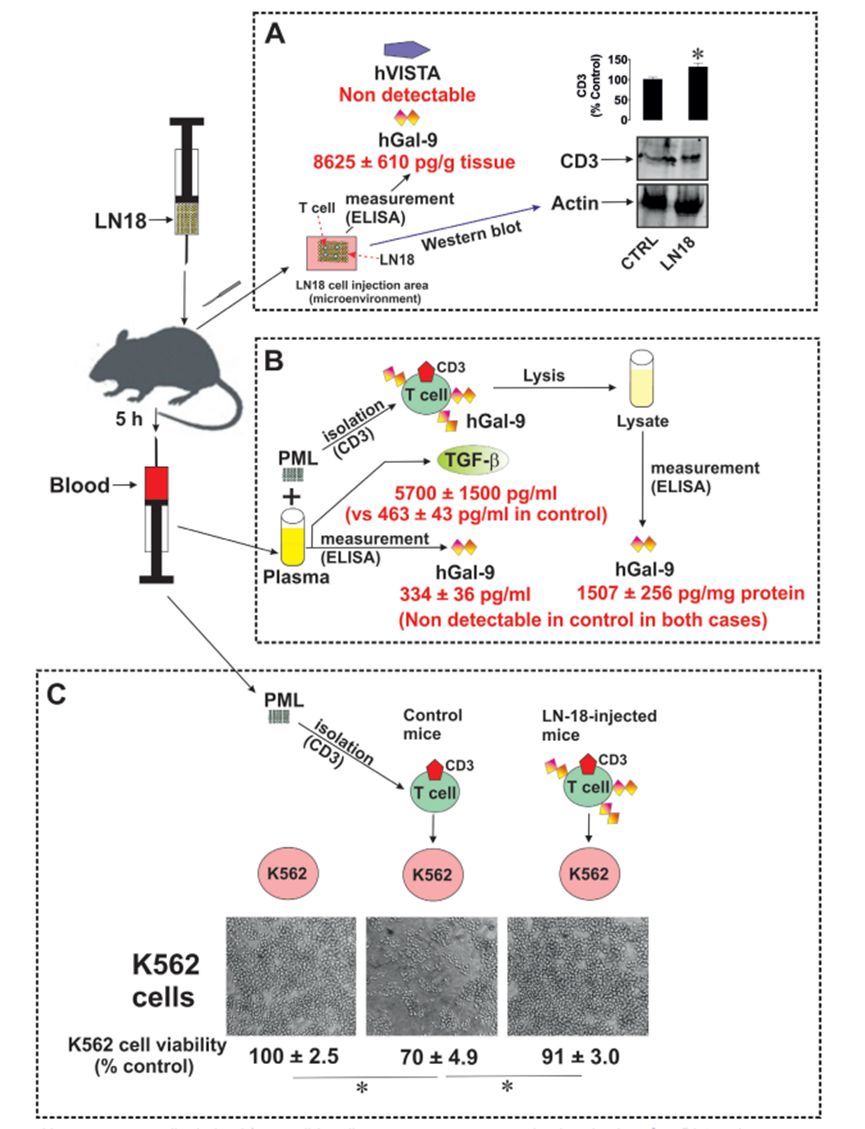
圖7、來源于實體惡性腫瘤的人癌細胞在體內分泌半乳糖凝集素-9。C57 BL16小鼠皮下注射致瘤性LN-18細胞(每只小鼠2 × 106個細胞)。然后在注射的微環境中測量CD3蛋白、人半乳糖凝集素-9和VISTA
結論
綜上所述,作者的研究結果首次表明,來自實體惡性腫瘤的人類癌細胞在與T細胞相互作用時能夠高度分泌半乳糖凝集素-9。半乳糖凝集素-9是通過轉運到細胞表面后的蛋白水解脫落以及與自噬和蛋白水解脫落相關的溶酶體分泌而分泌的。當非惡性細胞(如角質形成細胞)暴露于T細胞時,這一策略也適用,從而解釋了在銀屑病發展過程中,角質形成細胞如何抑制T細胞,從而抑制攻擊角質形成細胞的T細胞,隨后是角質形成細胞的增殖。當暴露于T細胞時,胎兒細胞不會分泌半乳糖凝集素-9,這表明胚胎細胞的目的只是抑制攻擊胚胎的T細胞,而不是抑制T細胞免疫本身。VISTA還通過抑制輔助性T細胞產生IL-2來增強半乳糖凝集素-9的免疫抑制作用,并可能進一步阻止細胞毒性T細胞的功能。這包括對PI-3K等信號通路的負面影響,影響BCL-XL抗凋亡蛋白的活性,以及如先前報道的通過改變其膜電位。因此,半乳糖凝集素-9本身及其產生相關的表達系統(如TGF-β/Smad3信號通路)可以被認為是多種癌癥免疫治療的特異性和高效靶點。
實驗方法
WB、ELISA、qPCR、細胞內鈣含量測定、On-cell Western、細胞活力測定
參考文獻
Schlichtner S, Yasinska IM, Lall GS, Berger SM, Ruggiero S, Cholewa D, Aliu N, Gibbs BF, Fasler-Kan E, Sumbayev VV. T lymphocytes induce human cancer cells derived from solid malignant tumors to secrete galectin-9 which facilitates immunosuppression in cooperation with other immune checkpoint proteins. J Immunother Cancer. 2023 Jan;11(1):e005714. doi: 10.1136/jitc-2022-005714. PMID: 36599470; PMCID: PMC9815087.