






Treg細胞來源的外泌體miR-2861-脊髓損傷治療的新靶點
血脊髓屏障(BSCB)是血液和脊髓實質之間的物理屏障。目前的證據表明,脊髓損傷后BSCB完整性的破壞可導致繼發性損傷,如脊髓水腫和過度炎癥反應。Treg細胞是有效的抗炎細胞,可抑制脊髓損傷后的神經炎癥反應,其在脊髓損傷后的浸潤表現出與BSCB自動修復相同的時空特征。然而,很少有研究評估Treg細胞與脊髓損傷之間的關系,強調BSCB的完整性。本研究探討Treg是否影響脊髓損傷后BSCB的恢復及其機制。我們證實脊髓損傷后同時發生血管生成和Treg細胞浸潤。此外,我們觀察到Treg細胞敲除和過表達對小鼠BSCB修復和運動功能的顯著影響。隨后,我們在體外證明了外泌體的存在和功能。此外,我們發現Treg細胞來源的外泌體包裹miR-2861,miR-2861調節血管緊密連接(TJs)蛋白的表達。熒光素酶報告基因實驗證實了miR-2861對IRAK1的負調控,一系列拯救實驗證實了IRAKI在調節BSCB中的生物學功能。總之,我們證明Treg細胞來源的外泌體可以包裝和遞送miR-2861并調節IRAK1的表達,從而影響小鼠脊髓損傷后BSCB的完整性和運動功能,這為脊髓損傷后的功能修復和限制炎癥提供了新的見解。本文于2023年10月發表于“Journal of Nanobiotechnology”(IF=10.2)上。
技術路線

結果
1)脊髓損傷后血脊髓屏障完整性的破壞和損傷部位Treg細胞的浸潤
在本研究中,我們利用CD31標記來跟蹤健康脊髓組織和損傷部位,以監測脊髓損傷后血管恢復和血脊髓屏障的任何破壞(圖1A)。我們發現,損傷部位的血管密度在第3天低于正常脊髓組織(1A, 1B)。第7天損傷部位與正常脊髓組織的血管密度無明顯差異(圖1A, 1B)。這些結果證實脊髓損傷后第3天損傷部位血管開始修復,并在第7天達到穩定狀態。然后將FITC-葡聚糖注射到脊髓損傷小鼠和正常小鼠體內,量化其熒光強度,觀察脊髓損傷后血脊髓屏障的狀態。我們發現,與正常組相比,脊髓損傷組損傷部位的熒光強度明顯增加(圖1C, 1D),這意味著BSCB受到嚴重損傷,導致SCI后FITC-葡聚糖滲透。對脊髓損傷后脊髓組織損傷區和非損傷區血管緊密連接相關蛋白進行免疫熒光染色,觀察血管緊密連接情況。我們的研究結果表明,與非損傷區域相比,ZO-1和Occludin與CD31在損傷區域的共定位顯著減少(圖1E-1H)。同時,western blotting顯示TJS相關蛋白的表達逐漸減少(圖1F)。這些結果證實了脊髓損傷后損傷區BSCB的完整性受到影響并逐漸加重。免疫熒光實驗顯示,脊髓損傷后損傷部位有顯著的Treg細胞浸潤(圖1H、1I)。據此,我們推測Treg細胞可能影響BSCB的修復。

2)Treg細胞促進脊髓損傷小鼠BSCB修復和運動功能恢復
為了探討Treg細胞對BSCB修復的影響,我們首先建立了Foxp3DTR轉基因小鼠,通過注射白喉毒素(DT)選擇性敲除Treg細胞并驗證敲除效率。將Treg細胞注射到Foxp3DTR轉基因小鼠的尾靜脈后,建立Treg細胞過表達小鼠模型。Foxp3DTR + DT組TJs蛋白與血管的共定位低于Foxp3DTR + PBS和Foxp3DTR + Treg組(圖2A、2B),而Foxp3DTR + Treg組TJs蛋白與血管的共定位明顯高于其他兩組(圖2A、2B)。這些結果表明敲除Treg細胞可以抑制SCI小鼠脊髓中TJs相關蛋白的表達。Treg細胞的過表達可以幫助TJs蛋白和血管更好地共定位,與Western blotting結果一致(圖2C)。為探討不同治療方法對脊髓損傷小鼠運動功能的影響,分別對三組脊髓損傷小鼠進行足跡分析。Foxp3DTR + DT組的足跡分析結果最差,其次是Foxp3DTR + PBS組,Foxp3DTR + Treg組(圖2D, 2E)。游泳試驗和Basso小鼠量表(BMS)行為分析顯示相似的結果(圖2F-2H)。這些結果證實了Treg細胞對脊髓損傷小鼠運動功能的影響。根據上述結果,我們認為損傷部位Treg細胞的積累可以促進脊髓損傷后小鼠BSCB的恢復,改善運動功能。

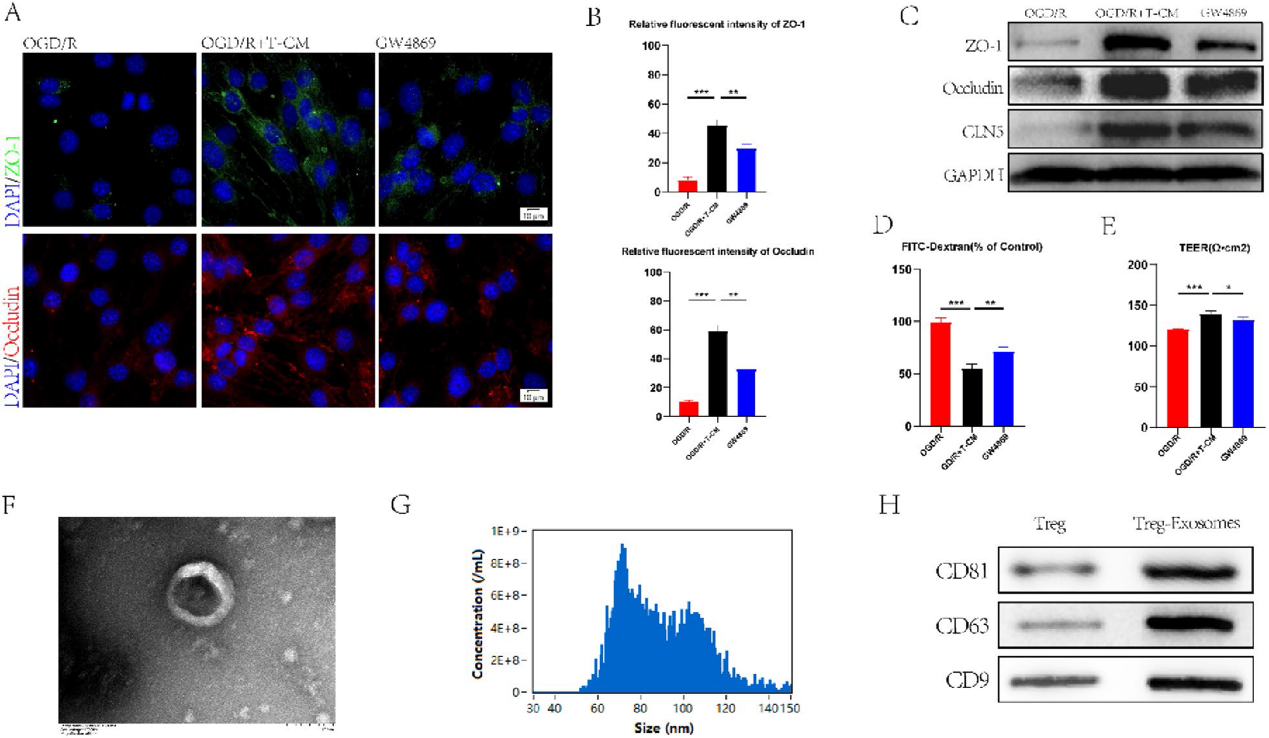
3)Treg來源的外泌體促進bEND.3細胞中緊密連接蛋白的表達
為了進一步探討Treg細胞對脊髓損傷小鼠BSCB的作用機制,我們采用氧-葡萄糖剝奪/再氧化(OGD/R)處理bEnd. 3細胞體外構建BSCB損傷模型,并進行免疫熒光染色。實驗結果表明,與OGD/R 處理的bEnd. 3細胞相比,T-CM處理組Occludin和ZO-1蛋白的表達明顯增加(圖3A, 3B)。細胞分泌的外泌體已被證明在細胞通訊中起作用。為了進一步探討T-CM的作用,用外泌體抑制劑GW4869處理與T-CM共培養的bEnd. 3細胞。GW4869的加入逆轉了T-CM對TJs蛋白表達的刺激作用(圖3A,3 B)。western blotting驗證了上述結果(圖3C)。內皮通透性和跨內皮電阻(TEER)被用來評估外泌體的治療效果(圖3D, 3E)。上述證據證實了Treg細胞可以通過分泌外泌體影響bEND. 3細胞中TJs蛋白表達的猜想。我們通過透射電子顯微鏡(TEM)和納米顆粒跟蹤分析(NTA)(圖3F, 3G)鑒定了Treg細胞來源的外泌體,并通過western blotting(圖3H)驗證了它們。
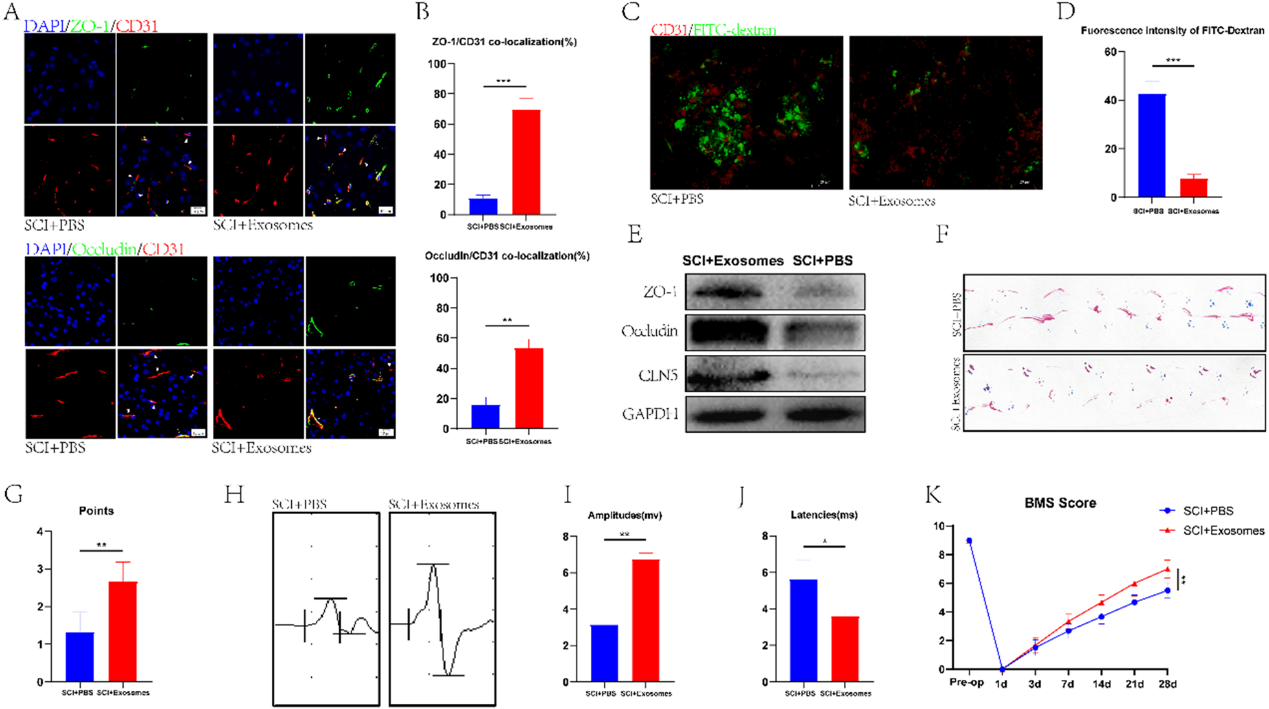
4)Treg來源的外泌體促進脊髓損傷小鼠的BSCB修復和運動功能恢復
為了探討Treg來源的外泌體(Treg-Exos)是否在SCI小鼠BSCB修復中發揮作用,我們將分離的外泌體注射到SCI小鼠體內,然后對小鼠脊髓進行免疫熒光染色。與SCI + PBS組相比,SCI +外泌體組Occludin和ZO-1與CD31的共定位更為明顯。(圖4A, 4B)。這些結果表明,注射外泌體后,SCI小鼠的BSCB修復更為顯著,與FITC-葡聚糖滲透實驗和western blot的結果相似(圖4C-4E)。我們通過多項行為分析比較了SCI +外泌體組與SCI + PBS組的運動功能,發現外泌體治療后,前者的運動功能較后者有明顯改善(圖4F-4K)。
5)外泌體在體外遞送miR-2861作用于bEND.3細胞以改善TJs蛋白表達
考慮到外泌體可以傳遞miRNA并發揮調控作用,我們從GEO數據庫中選擇了來自Treg細胞和源自Treg的外泌體的TOP20 miRNA,經交叉后獲得5個miRNA,其中選擇了miR-2861進行進一步研究。在確定miR-2861在外泌體中的存在后,我們試圖確認外泌體中的miR-2861是否可以在BSCB恢復中發揮關鍵的生物學作用。因此,我們通過慢病毒轉染敲低和過表達miR-2861,并分別構建相應的陰性對照(miR-2861KD、miR-NCKD、miR-2861OE和miR-NCOE)。通過qRT-PCR驗證轉染效率(圖5A)。為了觀察miR-2861在細胞水平上的作用,我們從慢病毒轉染的Treg細胞模型中提取外泌體(miR-NCKD-Exos, miR-2861KD-Exos, miR-NCOE-Exos和miR-2861OE-Exos),并將外泌體與OGD/ R處理的bEnd. 3細胞共培養。我們發現miR-2861在miR-2861KD-Exos組中的表達明顯低于miR-NCKD-Exos組。此外,miR-2861在miR-2861OE-Exos組中的表達明顯高于miR-NCOE-Exos組(圖5B)。在bEnd. 3個細胞中,各組miR-2861的表達趨勢相同,這與外泌體組miR-2861的表達水平相對應(圖5C)。免疫熒光染色顯示,miR-2861KD-Exos組Occludin和ZO-1的熒光強度明顯低于miR-NCKD-Exos組。在miR-2861OE-Exos組中觀察到相反的結果(圖5D,5 E),與WB(圖5F)一致。這些結果表明miR-2861的敲低和過表達可能會降低和增加TJs蛋白的表達。我們還觀察了bEND. 3細胞的滲透性和TEER。發現miR-2861KD-Exos組通透性增加,miR-2861OE-Exos組通透性降低(圖5G)。miR-2861KD-Exos組TEER顯著降低,miR-2861OE-Exos組TEER升高(圖5H)。以上結果有力地證明了miR-2861是一種促進BSCB修復的生物介質,探索其潛在機制具有重要意義。

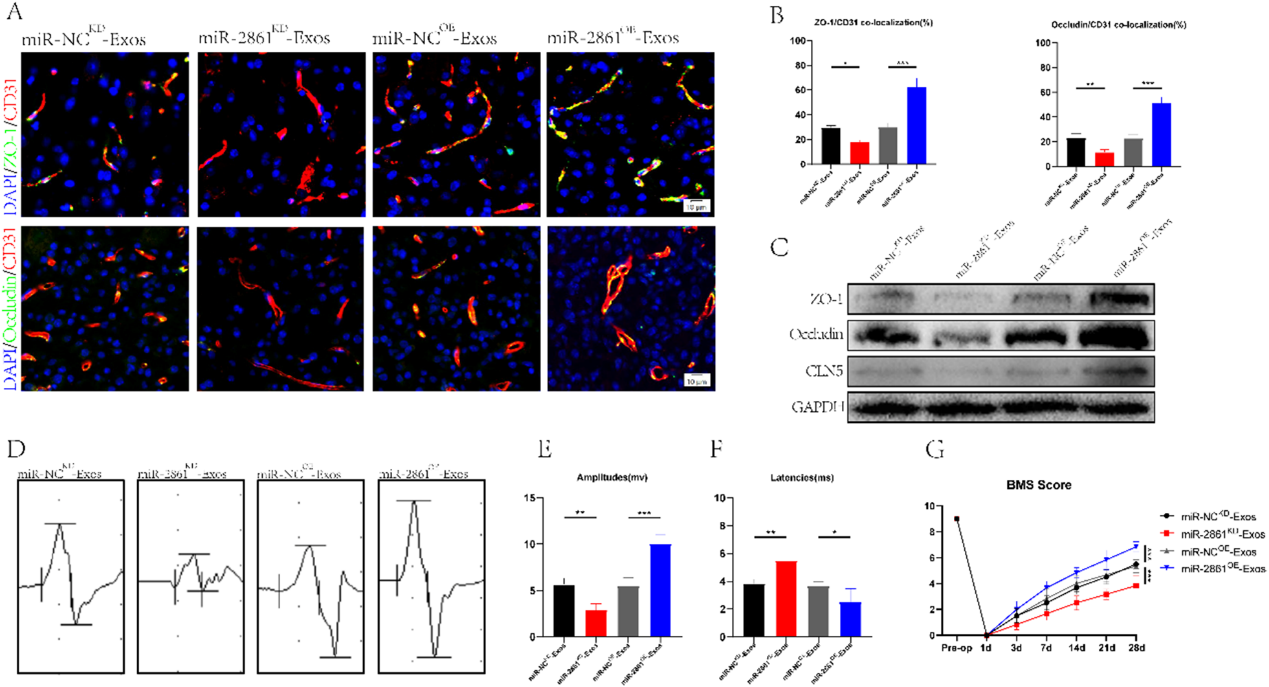
6)miR-2861在體內修復BSCB并增加運動功能
在驗證了miR-2861在細胞水平上的作用后,我們試圖闡明miR-2861在SCI小鼠體內的功能。我們將外泌體分為miR2861KD-Exos、miR-NCKD-Exos、miR-2861OE-Exos和miR-NCOE-Exos,并將其注射到SCI小鼠體內。我們發現miR-2861KD-Exos組TJs蛋白和血管的共定位強度明顯低于miR-NCKD-Exos組,而miR-2861OE-Exos組TJs蛋白和血管的共定位強度高于miR-NCOE-Exos組(圖6A, 6B)。這一發現得到了Western blot結果的支持(圖6C)。采用行為分析實驗觀察脊髓損傷小鼠運動功能的改善。MEP和BMS評分分析顯示,低表達miR-2861不利于運動功能的恢復,而高表達miR-2861可以增強運動功能(圖6D-6G)。總之,我們證實miR-2861可以通過外泌體運輸,調節BSCB的修復,從而改善運動功能。
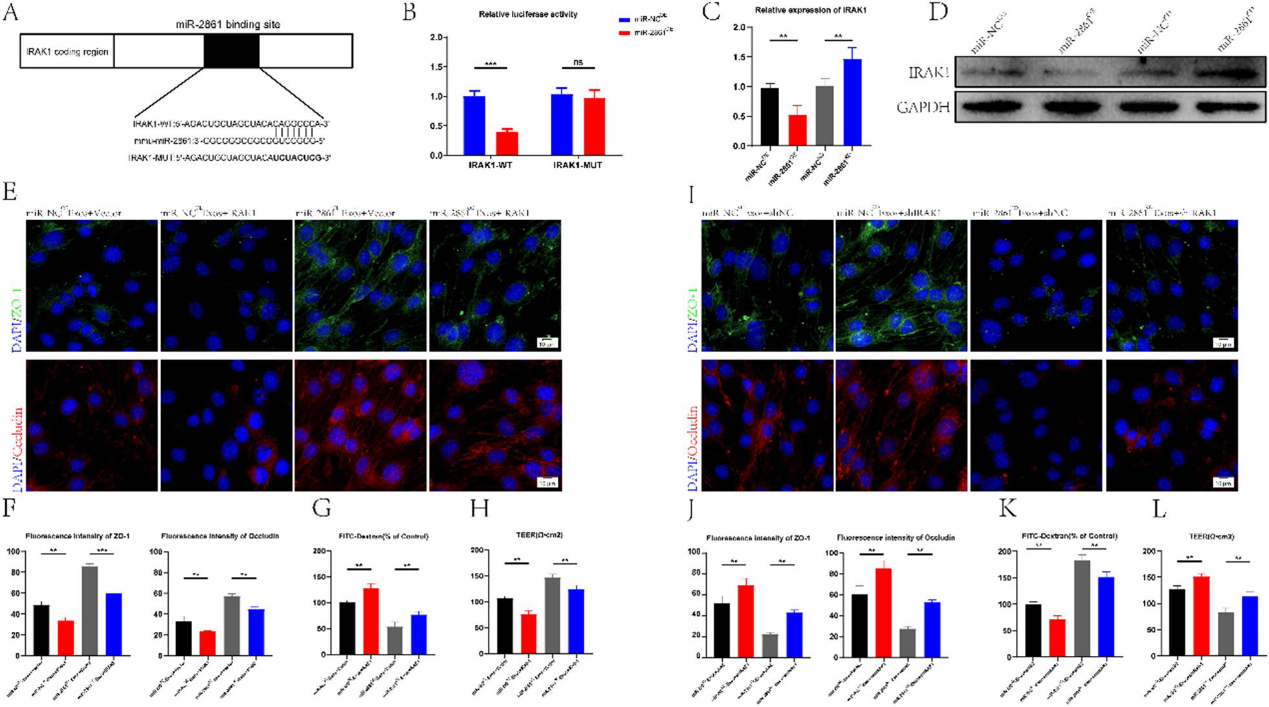
7)miR-2861通過抑制IRAK1表達促進BSCB修復
為了進一步探索miR-2861的具體機制,我們利用在線數據庫對miR-2861的潛在靶基因進行了分析和預測。其中,IRAK1與炎癥有關,其3 ' UTR可能被miR-2861識別并結合。為了驗證我們的猜想,基于IRAK1潛在結合位點構建IRAK1野生型(WT)和突變型(MUT) 3 ' -UTR序列(圖7A),并與miR-2861序列共轉染到bEnd. 3細胞。熒光素酶報告基因分析顯示,當WT-IRAK1-3'UTR和miR-2861OE組共轉染時,熒光素酶活性顯著低于WT-IRAK1-3'UTR和miR-NCOE組。然而,當MUT-IRAK1-3'UTR和miR-2861OE共轉染時,熒光素酶活性與對照組沒有顯著差異(圖7B),這表明IRAK1的3'UTR可以特異性結合miR-2861。我們進一步通過qRT-PCR和Western blot驗證了miR-2861可以負調控IRAK1(圖7C, 7D)。我們設計功能缺失和功能獲得實驗來進一步研究miR-2861與IRAK1之間的關系以及IRAK1對BSCB的影響。用miR-NCOE-Exos和miR-2861OE-Exos處理過表達IRAK1的bEnd. 3細胞,敲除IRAK1后,用miR-NCKD-Exos和miR-2861KD-Exos處理bEnd. 3細胞。結果表明,IRAK1過表達逆轉了miR-NCOE-Exos和miR- 2861OE-Exos誘導的TJs蛋白高表達(圖7E, 7F)。IRAK1過表達還逆轉了外泌體誘導的FITC-葡聚糖通透性降低和TEER升高(圖7G, 7H)。同樣,我們發現,敲低IRAK1可消除miR-NCKD-Exos或miR-2861KD-Exos對TJs的有害作用(圖7I,7 J)。FITC-葡聚糖通透性和TEER值產生相似的結果(圖7K, 7L)。以上數據提示miR-2681可以靶向IRAK1影響BSCB完整性。
結論
本研究發現脊髓損傷后損傷部位有Treg細胞浸潤。Treg細胞分泌的外泌體可促進BSCB的修復,降低其通透性,從而減輕浸潤引起的興奮損傷,促進運動功能的恢復。總之,我們的發現揭示了治療脊髓損傷的新靶點。
實驗方法
外泌體分離和鑒定,跨膜電阻測量,滲透率檢測,體內滲透性測定,游泳測試,BMS行為學分析,電生理檢測,WB,qRT-PCR,免疫熒光,熒光素酶報告試驗。
參考文獻
Kong G, Xiong W, Li C, Xiao C, Wang S, Li W, Chen X, Wang J, Chen S, Zhang Y, Gu J, Fan J, Jin Z. Treg cells-derived exosomes promote blood-spinal cord barrier repair and motor function recovery after spinal cord injury by delivering miR-2861. J Nanobiotechnology. 2023 Oct 4;21(1):364. doi: 10.1186/s12951-023-02089-6.