






天然多酚激活并增強GPX4以減輕淀粉樣蛋白β誘導的阿爾茨海默病鐵死亡

阿爾茨海默病(AD)是一種常見的神經退行性疾病,也是最常見的癡呆形式,其特征是記憶力和認知功能的進行性下降。盡管研究取得了重大進展,但對其病因的了解仍然有限。鐵死亡是一種鐵調節的細胞死亡方式,近年來因其參與多種疾病的發生發展而在病因學研究和治療方面受到關注。細胞內鐵水平是鐵死亡的關鍵因素。鐵還作為特異性脂氧合酶的輔因子促進酶促脂質過氧化。然而,啟動鐵死亡的鐵濃度閾值仍不明確。線粒體功能障礙、脂質過氧化以及抗氧化酶谷胱甘肽過氧化物酶4(GPX4)活性和水平的降低與鐵死亡相關。雖然鐵死亡正在被探索作為一種消滅腫瘤的治療途徑,但已發現它在多種神經系統疾病中發揮拮抗作用,包括AD。目前還需要開發同時靶向鐵死亡和AD的多功能分子,并協同增強和激活包括GPX4在內的天然抗氧化機制,以減輕AD中鐵死亡的復雜病理。該研究發表在《Chemical Science》,IF:9.825。
技術路線

圖1 機制圖

主要研究結果
1. 多酚(PPs)的鐵結合作用
鐵死亡是一種鐵依賴的細胞死亡方式,鐵螯合劑被認為是一種有前景的鐵死亡抑制劑。評估了一組主要含有兒茶酚和沒食子酸酯部分的PPs的鐵螯合能力。篩選的PPs包括沒食子酸(GA)、表沒食子兒茶素沒食子酸酯(EGCG)、鞣酸(TA)、鞣花酸(EA)、染料木黃酮(GEN)、丁香酸(SA)、沒食子酸甲酯(MeG)、4-羥基黃酮(4-HF)和2-羥基黃酮(2-HF)(圖1B)。在HEPES緩沖液中使用紫外-可見吸收光譜測定了這些PPs與Fe3+的相互作用強度。在加入FeCl3后,PPs的紫外-可見吸收光譜顯示吸光度增加,其中TA增加最大(圖2A)。將數據擬合為Benesi-Hildebrand方程,以確定結合常數(KB)。TA含有5個沒食子酸,表現出最高的鐵螯合能力,其KB值為6.64 ×105 M?1,而EGCG和GA的KB值分別為5.1×105 M?1和4.2×103 M?1(圖2B)。研究者發現MeG,GEN,EA和2-HF的結合常數遠低于TA。
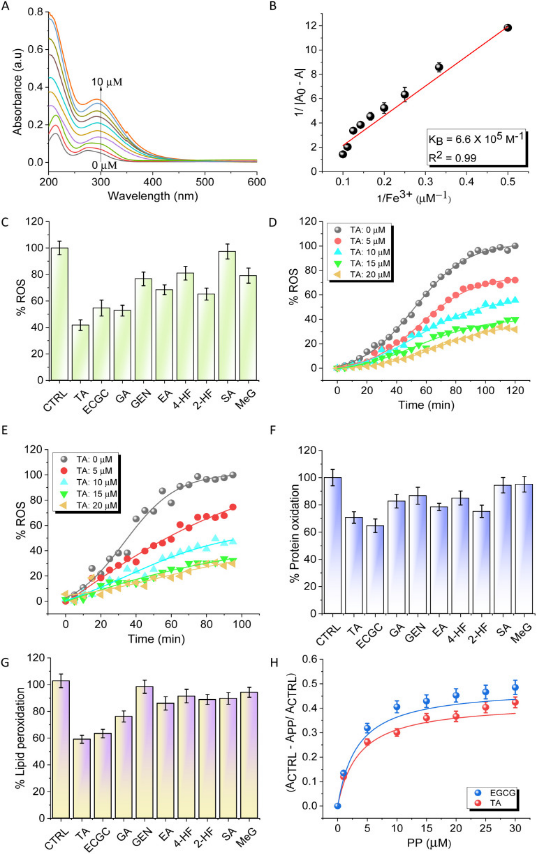
圖2 PPs的鐵螯合和抗氧化研究
2. 抗氧化作用和抑制脂質過氧化
采用α-二苯基-β-苦酰基肼(DPPH)、ABTS 和抗壞血酸鐵實驗評價強鐵螯合劑TA和EGCG以及GA的抗氧化活性。通過在PBS緩沖液和FeCl3中使用鐵抗壞血酸測定來評估PPs對氧化還原沉默Fe3+和猝滅過量ROS的能力(圖2C)。TA在452nm處以濃度依賴性的方式降低了發射,表明它能捕獲Fe3+并穩定氧化還原休眠狀態,并清除自由基(圖2D)。在最高濃度20μM時,TA對ROS的猝滅率為80%。有趣的是,在Aβ42存在的情況下進行的模擬AD條件的類似實驗表明,TA可以有效結合并阻止Fe3+氧化還原過程,從而猝滅ROS的產生(圖2E)。研究者使用Fe2++H2O2氧化還原體系來評估PPs對牛血清白蛋白氧化的保護作用,該體系在原位產生ROS,并模擬類似鐵死亡的鐵介導的氧化應激環境。TA和EGCG處理分別減少了30%和35%的蛋白質氧化(圖2F)。
接下來,研究者使用α-磷脂酰膽堿作為模型脂質和Fe2++H2O2氧化還原體系進行了脂質過氧化測定,以測量鐵死亡條件下的脂質過氧化程度。以Fe2++H2O2對α-磷脂酰膽堿的過氧化作用作為對照,發現TA和EGCG對脂質過氧化作用的抑制率分別為42%和40%(圖2G)。在TA和EGCG存在的情況下,脂質過氧化的濃度依賴性圖顯示了它們出色的過氧化抑制活性,這可以歸因于它們的協同鐵螯合和自由基捕獲機制(圖2H)。
3. 鐵非依賴性和依賴性Aβ42和tau蛋白聚集的調節
研究者評估了PPs調節Aβ42鐵非依賴性和依賴性聚集傾向的能力聚集依賴性硫黃素T(ThT)熒光數據顯示,在所有PPs中,EGCG和TA表現出優越的活性,在濃度為20μM時,聚集抑制率分別為44%和35%(圖3A)。Aβ42的聚集滯后時間(Tlag)為9.2h,一級聚集速率常數(κ)為0.51 h?1。TA和EGCG濃度的增加增加了Tlag,降低了κ,表明Aβ42聚集顯著延遲(圖3B)。當Aβ42∶PP為1∶5時,EGCG的抑制率(95%)明顯高于TA(82%)。由于鐵穩態在AD病因學中的關鍵作用,研究者研究了鐵對Aβ42聚集的影響。與原始Aβ42相比,Fe3+ 1:1的Aβ42使Tlag下降至8.72 h,而κ從0.51 h?1增加至0.72 h?1(圖3C)。在1:5比例下,TA和EGCG的Tlag值分別為10.12 h和10.9 h,κ值分別為0.20 h?1和0.17 h?1(圖3D)。
使用ThT熒光動力學監測PPs調節花生四烯酸誘導的tau聚集的能力(圖3E)。EGCG和TA在濃度為20μM時,對細胞的聚集抑制率分別為53%和46%。TA和EGCG表現出濃度依賴性的活性,在1:5的比例下,抑制率最高,分別為70%和81%,在2.5小時之后,沒有觀察到ThT熒光進一步增加(圖3F)。

圖3 篩選PPs抑制Aβ的能力42并使用ThT測定監測tau聚集
4. GPX4的激活及其RSL3誘導的抑制的逆轉
GPX4缺乏增加了細胞對鐵死亡的易感性。GPX4有兩個不同的位點:底物結合位點和變構位點。與抑制劑位點結合的小分子可作為抑制劑,而與激活位點結合預計可增強GPX4活性(圖4A)。TA與GPX4的激活位點結合,而EGCG與GPX4的激活位點或抑制位點不一致。研究者對GPX4抑制劑RSL3和GPX4激活劑PKUMDL-LC-102進行了對接研究(圖4A)。結果表明,RSL3與抑制位點結合,而PKUMDL-LC-102與激活位點結合,類似于TA。在變構激活位點,GPX4激活劑PKUMDL-LC-102被酸性殘基D21和D23以及極性殘基V98、F100和M102包圍(圖4A)。
使用GPX4抑制劑篩選試劑盒評估TA對GPX4活性的影響。在該實驗中,研究者利用GPX4對NADPH的消耗來監測酶的活性。GPX4與RSL3孵育60分鐘可抑制GPX4活性22%。然而,在20μM的濃度下,TA和EGCG能夠分別逆轉RSL3誘導的抑制作用12.3%和9.5%(圖4B)。TA通過結合GPX4的激活位點來增加GPX4的活性。通過監測RSL3對GPX4活性的濃度依賴性抑制,研究者計算出了RSL3的半最大抑制濃度(IC50)值,結果為17.17±0.17μM(圖4C,D)。RSL3的IC50值是在TA存在的情況下獨立計算的。在反應混合物中加入20 μM的TA將RSL3的IC50值從17.17±0.17μM調節到29.98±1.09μM,表明TA可以降低RSL3誘導的GPX4抑制(圖4E,F)。EGCG在RSL3誘導的GPX4抑制后顯示出相對較低的恢復內在酶活性的能力,將IC50值改變為21.01±0.47μM(圖5B)。對接結果表明TA具有與GPX4激活位點結合的能力。再加上TA可以有效逆轉RSL3誘導的GPX4抑制,因此研究者研究TA是否可以直接作為GPX4激活劑發揮作用。研究者的數據表明,用TA處理GPX4導致酶活性的濃度依賴性增加。在100 μM TA濃度下,觀察到GPX4活性增強了19%(圖4G)。這些結果表明,TA不僅可以從RSL3誘導的抑制中恢復GPX4的活性,而且還可以增強其內在活性。所有體外實驗的綜合分析確定TA是最有希望的抗鐵死亡候選藥物。這主要是因為它能夠激活GPX4并伴有多功能活性。相比之下,EGCG顯示出更好的抗淀粉樣蛋白和tau蛋白特性。
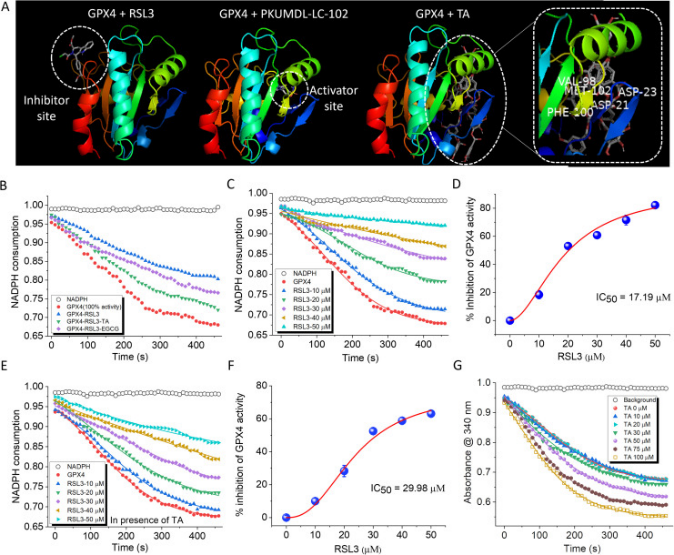
圖4 GPX4在小分子與抑制劑和激活劑位點結合的情況下的酶活性
5. 挽救鐵死亡誘導的細胞死亡
培養48h后,采用噻唑藍(MTT)法檢測鉛PPs對SH-SY5Y神經元細胞活力的影響。為了確定TA和EGCG抑制鐵死亡的能力,研究者將SH-SY5Y細胞分為RSL3單獨處理組和TA和EGCG聯合處理組。培養48h后,RSL3處理組細胞存活率為46%,而未處理組細胞存活率為100%。值得注意的是,TA和EGCG以濃度依賴性的方式拯救神經元細胞,在濃度為20μM時,細胞存活率分別達到84%和62%(圖5A,B)。研究者進行了TA與Ferrostatin-1和DFO的比較研究,以研究RSL3誘導的鐵死亡的拯救能力。Ferrostatin-1在10μM的低濃度下幾乎能100%抑制鐵死亡。TA挽救RSL3誘導的細胞死亡的效果與較低濃度的Ferrostatin-1相當。另一方面,盡管DFO具有比TA更強的鐵螯合能力,但在一定濃度下,DFO并未表現出顯著的拯救鐵死亡損傷的作用。采用免疫熒光法檢測RSL3單獨處理以及TA和EGCG聯合處理后細胞中GPX4的水平。與健康細胞相比,RSL3處理24 h可將GPX4水平降低至83%。值得注意的是,TA和EGCG處理使RSL3處理的細胞中的GPX4水平分別恢復至96%和91%,表明細胞從鐵死亡中恢復(圖5D)。此外,為了驗證這些結果,研究者使用了蛋白質印跡分析來定量GPX4的水平。與對照細胞相比,RSL3處理后GPX4水平降低至73%(圖5E、F)。值得注意的是,經TA治療后GPX4水平完全恢復。
LIP升高是鐵死亡的標志,可催化脂質過氧化并導致細胞死亡。用鈣黃綠素-乙酰氧基甲酯染色檢測RSL3單獨處理、TA和EGCG聯合處理的細胞LIP。鈣黃綠素-乙酰氧基甲酯在細胞內被酯酶裂解,在細胞質中產生鈣黃綠素熒光團,表現出高熒光。然而,當鈣黃綠素與LIP螯合時,其熒光被猝滅,從而可以估算游離LIP水平。與健康細胞相比,RSL3處理的細胞顯示細胞內Fe水平增加,相應的鈣黃綠素熒光降低至67%(圖5G-K)。值得注意的是,50μM TA和EGCG處理降低了LIP水平,導致鈣黃綠素熒光分別增加約95%和100%,與對照細胞相似(圖5L)。這種作用呈劑量依賴性,表明TA和EGCG能夠隔離游離Fe并使其失活。鐵死亡觸發脂質過氧化,增加ROS生成,導致細胞內線粒體ROS水平升高和線粒體膜電位(MMP)下降,導致線粒體碎片化。研究者使用MitoSOX探針監測細胞中線粒體ROS水平。RSL3處理使線粒體ROS水平增加到148%,與健康細胞相比。相反,TA和EGCG分別將RSL3處理的細胞中的ROS水平降低至95%和91%,表明它們有能力預防鐵死亡ROS介導的線粒體損傷(圖5M)。通過探針檢測MMP的變化,證實RSL3誘導的鐵死亡和PP處理的逆轉。RSL3處理細胞后,MMP下降至76%,與正常細胞比較差異有統計學意義。然而,TA和EGCG處理分別將MMP恢復至93%和95%,表明對功能性線粒體損傷有預防作用(圖5N)。采用Mito-TG.45染色法檢測TA和EGCG對鐵死亡細胞線粒體結構損傷的保護作用采用活細胞熒光顯微鏡觀察RSL3單獨處理或TA和EGCG聯合處理的細胞。值得注意的是,經TA和EGCG處理的細胞保持了健康的管狀結構,支持這些PPs對鐵死亡線粒體損傷的保護作用,ROS和MMP研究證明了這一點(圖5R,S)。

圖5 細胞研究表明TA和EGCG能夠從鐵死亡和Aβ中拯救神經元細胞42-誘導毒性
6. 從淀粉樣蛋白誘導的鐵死亡中拯救神經元細胞
TA和EGCG調節Aβ42聚集的有效性促使研究者研究它們拯救神經細胞免受Aβ42誘導的毒性的能力。分別用Aβ42、TA和EGCG處理SH-SY5Y細胞。Aβ42處理細胞30h后,細胞存活率為34.8%,與未處理的對照組相比,差異有統計學意義。然而,在PPs存在的情況下,研究者觀察到Aβ42處理的神經元細胞以劑量依賴性的方式得到挽救,最高濃度的TA和EGCG分別使細胞存活率提高到63%和69%(圖5T)。在AD的鐵死亡背景下,鐵加速毒性淀粉樣蛋白(Aβ42-Fe)纖維的積累,產生活性氧,導致膜損傷和脂質過氧化。研究者使用MTT法檢測TA和EGCG調節毒性Aβ42 + Fe聚集和拯救神經元細胞免于Aβ42 + Fe誘導的鐵死亡的能力。Aβ42: Fe(1: 5)處理組細胞存活率為17%,對照組細胞存活率為100%。然而,用TA和EGCG處理后,Aβ42 + Fe處理的細胞存活率分別提高到74%和77%,呈劑量依賴性(圖5U,V)。將細胞分為Aβ42 + Fe組和TA + EGCG組,DCFDA法檢測細胞內ROS水平;Aβ42 + Fe(1:5)培養6h后,細胞內ROS由未處理組的31%增加至100%。然而,TA和EGCG處理后,Aβ42 + Fe處理細胞中的ROS水平分別降低至36%和38%,表明它們具有復合體和使Fe氧化還原失去活性、調節Aβ42 - Fe fenton型反應并最終阻止ROS生成的能力(圖50)。這些細胞研究有力地支持TA和EGCG能夠減輕RSL3誘導的鐵死亡和Aβ42 - Fe相關毒性,使它們成為協同靶向鐵死亡和AD的潛在候選藥物。結合ThT研究,研究者評估了TA保護神經元細胞免受tau誘導的毒性的能力。在AA誘導的tau聚集物存在下,與健康細胞相比,細胞活力降低至47%(圖5W)。TA可以劑量依賴性地增加細胞活力,將細胞活力提高到100%。
為了證實Aβ42與鐵死亡之間的關系,研究者采用免疫熒光法檢測Aβ42 + Fe單獨孵育以及TA和EGCG共同孵育的SH-SY5Y神經元細胞株中GPX4的水平。與健康細胞相比,Aβ42 + Fe(1:5)處理細胞30小時顯著降低GPX4水平至65%(圖6)。據研究者所知,這是第一個在鐵死亡背景下的神經細胞中Aβ42 + Fe和GPX4水平直接相關的報告,通過直接定量蛋白水平的免疫熒光測定確定。與未處理的對照細胞相比,TA顯著增加了Aβ42 + Fe處理的細胞GPX4水平(147%),表明TA可以在AD病理條件下提高GPX4水平。
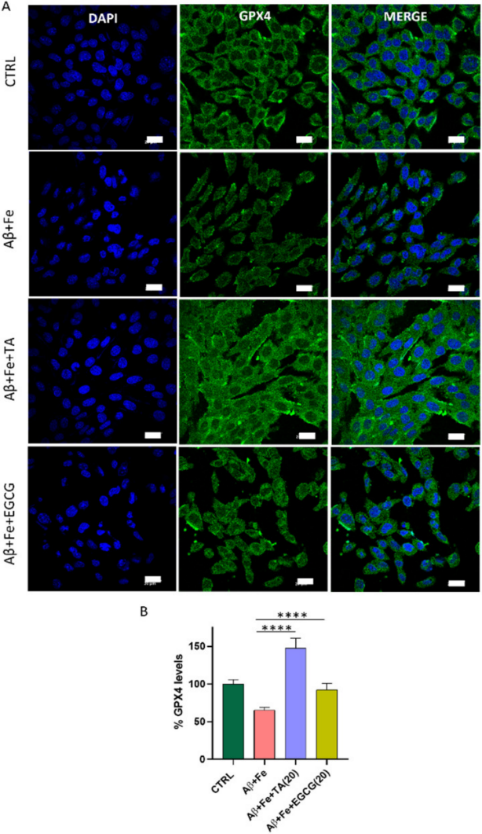
圖6 通過免疫熒光(IF)測定進行蛋白質定量
這些發現促使研究者在神經元細胞中檢測TA作為GPX4增強子的作用。研究者觀察到,與未處理的細胞相比,TA處理的細胞內GPX4水平增加到192%(圖7A,B)。TA不僅增強GPX4活性,而且增加其在細胞中的水平。蛋白質印跡檢測GPX4蛋白表達水平。結果顯示,與未處理的細胞相比,TA處理使GPX4水平增加到127%(圖7C,D)。基于這些發現,研究者檢測了抗氧化主要調節因子核因子e2相關因子2(Nrf2)的變化,已知Nrf2可直接和間接調節GPX4及其相關抗氧化蛋白46雖然有報道表明TA能夠激活Nrf2通路,但研究者評估了在神經細胞中TA處理后Nrf2的激活。研究表明,TA處理導致SH-SY5Y細胞中Nrf2積累和核轉位。與未處理的細胞相比,細胞內Nrf2水平增加到148%,并且使用TA處理后,來自細胞質的Nrf2蛋白發生了2倍的核易位(圖7E-G)。因此,研究者推測TA可能通過Nrf2-GPX4軸提高GPX4水平。

圖7 通過IF測定和蛋白質印跡進行蛋白質定量
結論
該研究發現天然的TA是一種高效的鐵死亡抑制劑。它通過鐵螯合、抑制脂質過氧化、拯救線粒體損傷和激活Nrf2-GPX4軸等鐵死亡的所有主要途徑發揮作用,從而提高它們的水平。同時,TA是一種有效的Aβ42和tau蛋白聚集的抑制劑,有效地調節劇毒鐵誘導的Aβ42聚集,減少氧化應激,對抗AD患者的鐵穩態失調。雖然GPX4蛋白的合成效率非常低且需要大量能量,但研究者證明了一種天然的多酚如何提供一種可行的解決方案,以協同調節鐵死亡和AD的多方面毒性,而GPX4的激活是關鍵機制之一。這可能鼓勵研究人員探索無毒的天然產物來解決問題,并強調GPX4在有效調節鐵死亡以及闡明其在AD中的作用的重要性。
實驗方法
ABTS抗氧化試驗,鐵抗壞血酸測定,硫黃素T(ThT)熒光檢測,細胞培養和MTT法檢測,ROS測定,神經細胞拯救實驗,溫滴定量熱法(ITC)測量,蛋白質印跡,免疫熒光
參考文獻
Baruah P, Moorthy H, Ramesh M, Padhi D, Govindaraju T. A natural polyphenol activates and enhances GPX4 to mitigate amyloid-β induced ferroptosis in Alzheimer's disease. Chem Sci. 2023 Aug 22;14(35):9427-9438. doi: 10.1039/d3sc02350h.