






LncRNA MGCG-膠質(zhì)母細(xì)胞瘤的治療靶點(diǎn)
膠質(zhì)母細(xì)胞瘤(GBM)是成人最常見(jiàn)的原發(fā)性惡性腦癌。lncRNAs在癌癥的發(fā)生發(fā)展中發(fā)揮著重要作用。為了闡明lncRNA在膠質(zhì)母細(xì)胞瘤發(fā)展中的作用,我們使用了來(lái)自GBM患者的三個(gè)新分離的腫瘤組織樣本和來(lái)自創(chuàng)傷患者的三個(gè)正常腦組織樣本,進(jìn)行了高通量RNA測(cè)序以獲得轉(zhuǎn)錄本。隨后,一種lncRNA MGCG在GBM中未被報(bào)道,被發(fā)現(xiàn)與患者預(yù)后相關(guān)。生物信息學(xué)分析結(jié)果顯示,MGCG與自噬相關(guān),且與自噬相關(guān)基因ATG2A的表達(dá)呈正相關(guān)。質(zhì)譜數(shù)據(jù)表明,hnRNPK蛋白是與MGCG相互作用的直接靶點(diǎn),MGCG/hnRNPK通過(guò)增強(qiáng)ATG2A的翻譯和自噬促進(jìn)GBM的發(fā)展。綜上所述,本研究表明MGCG具有促進(jìn)GBM發(fā)展的潛力,可能成為腫瘤分子診斷和治療的候選藥物。本研究于2023年7月發(fā)表于“Cell Death and Disease”(IF=9.0)上。
技術(shù)路線
結(jié)果
1)MGCG在GBM中高水平表達(dá),與預(yù)后不良相關(guān)
為了獲得lncRNAs和mRNA的表達(dá)譜,我們獲得3個(gè)GBM組織樣本和3個(gè)正常腦組織樣本,并進(jìn)行高通量RNA測(cè)序(GSE153692)。RNA測(cè)序結(jié)果顯示,MGCG在GBM組織中的表達(dá)明顯高于正常組織(圖1A)。接下來(lái),我們通過(guò)RT-qPCR驗(yàn)證了MGCG在22個(gè)正常腦組織樣本(顱腦外傷患者)和47個(gè)GBM組織樣本中的表達(dá),結(jié)果表明MGCG在GBM組織中的表達(dá)水平明顯高于正常組織(圖1B)。RT-qPCR結(jié)果顯示,MGCG在GBM細(xì)胞中的表達(dá)明顯高于HEB細(xì)胞(圖1C)。為了確定MGCG的細(xì)胞定位,我們對(duì)細(xì)胞核和細(xì)胞質(zhì)RNA進(jìn)行了RT-qPCR,數(shù)據(jù)顯示MGCG優(yōu)先位于細(xì)胞核中(圖1D)。接下來(lái),我們?cè)贚N229細(xì)胞中進(jìn)行RNA FISH驗(yàn)證這些結(jié)果(圖1 E)。RNA FISH結(jié)果顯示,MGCG在GBM中與正常組織相比上調(diào)(圖1F)。我們?cè)u(píng)估了MGCG表達(dá)與GBM患者預(yù)后之間的相關(guān)性。Kaplan-Meier分析和MGCG表達(dá)與臨床患者生存期的相關(guān)性分析結(jié)果顯示,MGCG表達(dá)較高的患者總體生存期較差(圖1G)。綜上所述,這些數(shù)據(jù)提示MGCG上調(diào)在GBM組織中普遍存在,且與預(yù)后不良相關(guān)。

2)MGCG促進(jìn)GBM的腫瘤進(jìn)展和自噬
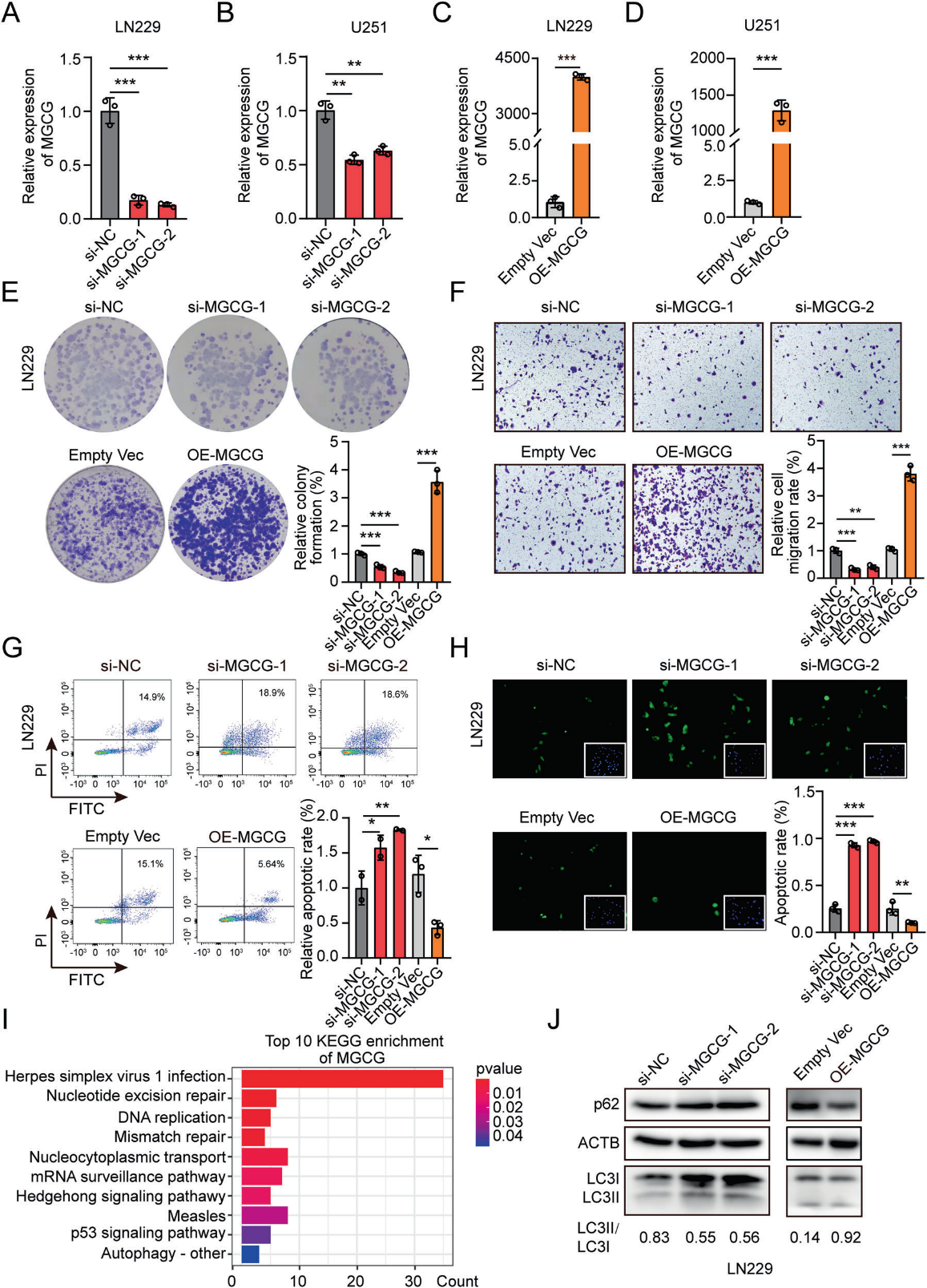
為了探索MGCG的生物學(xué)功能,我們?cè)O(shè)計(jì)了三種獨(dú)立的siRNA來(lái)對(duì)抗MGCG,我們選擇了兩種siRNA來(lái)有效地抑制MGCG在LN229和U251細(xì)胞中的表達(dá)(圖2A, 2B)。我們還構(gòu)建了一個(gè)過(guò)表達(dá)質(zhì)粒,并成功地在LN229和U251細(xì)胞中過(guò)表達(dá)MGCG(圖2C, 2D)。集落形成實(shí)驗(yàn)的結(jié)果表明,MGCG的缺失顯著抑制了細(xì)胞增殖。而MGCG的過(guò)表達(dá)促進(jìn)了細(xì)胞活力(圖2E)。我們?cè)赥ranswell實(shí)驗(yàn)中證實(shí),MGCG的下調(diào)顯著降低了細(xì)胞的遷移能力;然而,MGCG過(guò)表達(dá)增加了細(xì)胞的遷移能力(圖2F)。此外,流式細(xì)胞術(shù)和TUNEL實(shí)驗(yàn)結(jié)果表明,MGCG的敲低誘導(dǎo)細(xì)胞凋亡,上調(diào)MGCG的表達(dá)抑制細(xì)胞凋亡(圖2G,2 H)。基于RNA測(cè)序數(shù)據(jù)研究MGCG在GBM中的作用,我們進(jìn)行了生物信息學(xué)分析,并證明MGCG與自噬有關(guān)(圖2I)。western blotting結(jié)果顯示,MGCG過(guò)表達(dá)增加了LC3-II結(jié)合和p62降解,表明MGCG誘導(dǎo)了GBM細(xì)胞的自噬(圖2J)。結(jié)果表明,MGCG的表達(dá)與GBM的惡性表型相關(guān),MGCG促進(jìn)了GBM的自噬。
3)MGCG通過(guò)調(diào)節(jié)ATG2A的表達(dá)促進(jìn)GBM的進(jìn)展
許多研究表明,自噬促進(jìn)GBM的發(fā)展。一致地,我們證明了自噬在過(guò)表達(dá)MGCG的GBM細(xì)胞中升高。ATG2A是一種與自噬相關(guān)的基因,我們已經(jīng)知道敲低ATG2A會(huì)抑制自噬,但對(duì)ATG2A的研究很少。RT-qPCR數(shù)據(jù)證明,在GBM組織臨床樣本中,MGCG的表達(dá)與ATG2A呈顯著正相關(guān)(圖3A)。接下來(lái),我們敲低了ATG2A,發(fā)現(xiàn)LN229細(xì)胞中LC3-II與LC3-I的蛋白水平比值降低,p62蛋白水平升高(圖3B)。為了研究MGCG是否調(diào)控ATG2A的表達(dá),我們敲低了MGCG,分別根據(jù)RT-qPCR和western blotting的數(shù)據(jù)證實(shí)了ATG2A mRNA和蛋白的表達(dá)水平降低(圖3C, 3D)。我們通過(guò)RT-qPCR驗(yàn)證了ATG2A在27個(gè)正常組織樣本和42個(gè)GBM組織樣本中的表達(dá),證實(shí)了ATG2A在GBM組織中的表達(dá)水平顯著升高(圖3E)。此外,根據(jù)TCGA數(shù)據(jù)庫(kù),ATG2A的低表達(dá)與患者總生存率的提高相關(guān)(圖3F)。此外,我們還研究了ATG2A對(duì)GBM進(jìn)展的影響。首先,我們通過(guò)RT-qPCR和western blot檢測(cè)ATG2A在GBM細(xì)胞中的敲除效率(圖3G-3J)。ATG2A的下調(diào)抑制了GBM細(xì)胞的增殖(圖3K)和遷移(圖3L),促進(jìn)了GBM細(xì)胞的凋亡(圖3M)。然后,我們將含有ATG2A siRNA的質(zhì)粒和過(guò)表達(dá)MGCG的質(zhì)粒共轉(zhuǎn)染到LN229和U251細(xì)胞中,我們證明了ATG2A的下調(diào)抑制了自噬,過(guò)表達(dá)MGCG恢復(fù)了自噬的抑制(圖3N)。敲低ATG2A可消除MGCG過(guò)表達(dá)對(duì)細(xì)胞增殖、遷移和凋亡的影響(圖3O-Q)。這些結(jié)果表明MGCG調(diào)節(jié)ATG2A的表達(dá),促進(jìn)細(xì)胞自噬。

4)MGCG直接與hnRNPK相互作用,促進(jìn)ATG2A的翻譯
為了進(jìn)一步探索MGCG在GBM中的作用機(jī)制,我們使用特異性生物素標(biāo)記的MGCG探針進(jìn)行了RNA下拉實(shí)驗(yàn)。與陰性探針相比,銀染色結(jié)果顯示了幾個(gè)額外的蛋白質(zhì)條帶,質(zhì)譜分析鑒定出hnRNPK是與MGCG結(jié)合的主要蛋白質(zhì)(圖4A,B)。RIP實(shí)驗(yàn)的數(shù)據(jù)證實(shí)了MGCG和hnRNPK之間的直接相互作用(圖4C)。FISH和IF數(shù)據(jù)顯示,MGCG和hnRNPK在GBM細(xì)胞中共定位(圖4D)。western blot分析結(jié)果顯示,MGCG的敲低或MGCG的過(guò)表達(dá)分別抑制或升高了GBM細(xì)胞中hnRNPK的蛋白水平(圖4E)。為了確定hnRNPK是否可以調(diào)節(jié)ATG2A的表達(dá),我們進(jìn)行了RT-qPCR,并證明了hnRNPK敲低后ATG2A的RNA水平?jīng)]有變化(圖4F)。western blotting結(jié)果顯示,當(dāng)hnRNPK被敲低時(shí),ATG2A蛋白水平下調(diào)(圖4G)。為了確定hnRNPK是否調(diào)控ATG2A在GBM中的翻譯,我們使用RIP證明hnRNPK與ATG2A mRNA結(jié)合(圖4H),RNA下拉結(jié)果表明hnRNPK與ATG2A mRNA之間存在相互作用(圖4I)。接下來(lái),我們進(jìn)行了IP分析,證實(shí)了hnRNPK在GBM中與EIF4B結(jié)合(圖4J)。我們通過(guò)far western blot進(jìn)一步證實(shí)了hnRNPK與EIF4B之間的相互作用(圖4K)。western blot結(jié)果顯示,敲低EIF4B抑制ATG2A蛋白水平;然而,在LN229細(xì)胞中,下調(diào)EIF4B后,ATG2A的RNA水平?jīng)]有變化(圖4L, 4M)。敲低hnRNPK降低ATG2A蛋白水平;然而,過(guò)表達(dá)MGCG消除了hnRNPK敲低的作用(圖4N)。這些結(jié)果表明,MGCG通過(guò)結(jié)合hnRNPK促進(jìn)了ATG2A的翻譯。

5)DNA低甲基化上調(diào)GBM中MGCG的表達(dá)
我們探討了MGCG在GBM中高表達(dá)的機(jī)制。首先,我們利用UCSC基因組生物信息學(xué)網(wǎng)站確定了MGCG中的CpG島(圖5A),數(shù)據(jù)表明MGCG表達(dá)與DNA甲基化之間存在潛在關(guān)系。利用甲基化分析軟件,我們?cè)贛GCG啟動(dòng)子區(qū)域檢測(cè)到一個(gè)CpG島。設(shè)計(jì)了兩組特異性引物來(lái)檢測(cè)該CpG島DNA甲基化水平的變化(圖5B)。接下來(lái),使用甲基化特異性PCR (MSP),我們證明GBM細(xì)胞的甲基化水平低于HEB細(xì)胞(圖5C)。為了進(jìn)一步探討MGCG中甲基化變化的機(jī)制,我們檢測(cè)了DNA甲基轉(zhuǎn)移酶(DNMT)在GBM細(xì)胞中的表達(dá)水平。RT-qPCR數(shù)據(jù)顯示,與HEB細(xì)胞相比,DNMT3B在GBM細(xì)胞中的表達(dá)下調(diào)(圖5D)。我們用DNMT3B特異性抑制劑DS-437處理LN229和U251細(xì)胞,發(fā)現(xiàn)MGCG在LN229和U251細(xì)胞中的表達(dá)增加,表明DNA去甲基化增加了MGCG的表達(dá)(圖5E)。DNMT3B過(guò)表達(dá)導(dǎo)致MGCG甲基化水平增加(圖5F)。過(guò)表達(dá)DNMT3B可顯著抑制MGCG在GBM細(xì)胞中的表達(dá)(圖5G)。這些發(fā)現(xiàn)表明DNMT3B促進(jìn)了MGCG在GBM細(xì)胞中的表達(dá)。

6)MGCG/hnRNPK/ATG2A軸促進(jìn)GBM的進(jìn)展
為了研究MGCG在體內(nèi)的作用,將2.5×105熒光素標(biāo)記的MGCG敲低或sh-NC轉(zhuǎn)導(dǎo)的GBM細(xì)胞注射到裸鼠體內(nèi),體內(nèi)腫瘤異種移植實(shí)驗(yàn)方案如圖6A所示。通過(guò)體內(nèi)生物發(fā)光成像追蹤腫瘤進(jìn)展,攜帶MGCG敲低的GBM細(xì)胞的異種移植物在腫瘤生長(zhǎng)方面表現(xiàn)出明顯的退行(圖6B,6C)。MGCG敲低導(dǎo)致小鼠存活率提高(圖6D)。RNA FISH數(shù)據(jù)顯示,MGCG敲低小鼠GBM細(xì)胞中MGCG表達(dá)降低(圖6E)。免疫組化(IHC)顯示,MGCG敲低后hnRNPK和ATG2A表達(dá)降低,cleaved caspase-3水平升高(圖6F),這些結(jié)果表明MGCG/hnRNPK/ATG2A促進(jìn)了GBM的進(jìn)展。

結(jié)論
我們發(fā)現(xiàn)了一種新的lncRNA-MGCG,作為一種致癌基因,促進(jìn)GBM的增殖和遷移。接下來(lái),我們驗(yàn)證了MGCG與自噬有關(guān)。我們證明MGCG直接與hnRNPK結(jié)合,并且MGCG/ hnRNPK促進(jìn)ATG2A的翻譯。因此,本研究揭示了lncRNA調(diào)控GBM生長(zhǎng)的機(jī)制,并確定了GBM治療的潛在治療藥物靶點(diǎn)。
實(shí)驗(yàn)方法
qRT-PCR,F(xiàn)ISH,克隆形成實(shí)驗(yàn),transwell,TUNEL實(shí)驗(yàn),流式,WB,F(xiàn)ar WB,IF,RNA pull down,RIP,IHC。
參考文獻(xiàn)
Chu F, Wu P, Mu M, Hu S, Niu C. MGCG regulates glioblastoma tumorigenicity via hnRNPK/ATG2A and promotes autophagy. Cell Death Dis. 2023 Jul 17;14(7):443. doi: 10.1038/s41419-023-05959-x.