






Cell熱點:大腦皮層類器官聯合多組學分析
要想充分發揮腦類器官研究人類發育的作用,就必須了解類器官是否精確復制了內源性細胞和分子事件,特別是因為類器官中細胞身份的獲得可能會受到異常代謝狀態的影響。本文展示了人類大腦皮層類器官發育的綜合單細胞轉錄組、表觀遺傳組和空間圖譜,包括超過61萬個細胞,從神經祖細胞的生成到分化的神經元和膠質亞型的產生。本研究表明,無論新陳代謝狀態如何,細胞多樣化過程都與內源性細胞密切相關,因此,可以利用類器官圖譜研究人類命運的決定。作者定義了器官發育過程中皮質細胞類型的縱向分子軌跡,確定了在細胞系建立過程中具有人類特異性作用的基因,并發現了人類胼胝體神經元的早期轉錄多樣性。這些發現驗證了人類類器官皮質發生綜合圖譜是研究人類皮質發育機制的首要資源。本文于2022年9月發表在《Cell》IF:64.5期刊上。
技術路線

主要實驗結果
1、人類皮質類器官發育的單細胞水平轉錄組、表觀基因組和空間圖譜
為研究人腦類器官中皮質發生的分子特征和可重復性,構建了一個縱向單細胞圖譜,包含類器官發育6個月期間的8個時間點,涵蓋從早期祖細胞擴增到后期星形膠質細胞產生的過程(圖1A),并在RNA、染色質和空間轉錄組學水平進行了分析。本文的轉錄組數據集包含本研究scRNA-seq檢測出的532,414個細胞,并結合了作者之前發表的數據集,其來自多個時間點的218,240個新鑒定出的細胞,共計83個單獨鑒定出的類器官,來自多個干細胞系和分化批次(圖1A-1B)。表觀基因組數據集包括38,017個核,包括單細胞ATAC-seq獲得的轉座酶可及染色質,并結合了作者之前發表的數據集,其包含體外培養1、3和6個月后類器官的11,551個新發現的細胞核,分別跨越了神經祖細胞的擴增、興奮性神經元多樣性的峰值以及星形膠質細胞和中間神經元的出現(圖1C)。此外,納入了通過SHARE-seq分析的42,810個細胞,SHARE-seq在從23日至3個月的4個時間點同時從同一單細胞采集了RNA-seq和ATAC-seq分析結果(圖1D)。最后,空間轉錄組數據集包含10個類器官,這些類器官在體外1、2和3個月時通過Slide-seqV2分析得到(圖2),涵蓋興奮性神經元的出現和擴張,這些神經元在體外3個月時類器官的多樣性最高。這些數據提供了人類皮質類器官發育的綜合多組學分子圖譜.
作者評估了這些細胞產生的縱向事件在所有類器官的發育過程中可重復性,通過計算每個采樣時間點的調整后互信息(AMI)得分,衡量了存在的細胞類型比例與單個起源的類器官之間的依賴關系(圖1E)。在各年齡段和細胞系中,大多數重復的 AMI 分數都與兩個已發表的內源性人類皮層數據集和一個新生成的胎兒數據集相當。并且總體而言,scRNA-seq和scATAC-seq定義的細胞類型之間存在良好的一致性,AMI評分表明,個體類器官之間的表觀遺傳變異性與轉錄變異性處于相同的低水平(圖1F)。表明這些隨著時間延伸的類器官數據也在多組學層面展現出了細胞分化的可重復性。
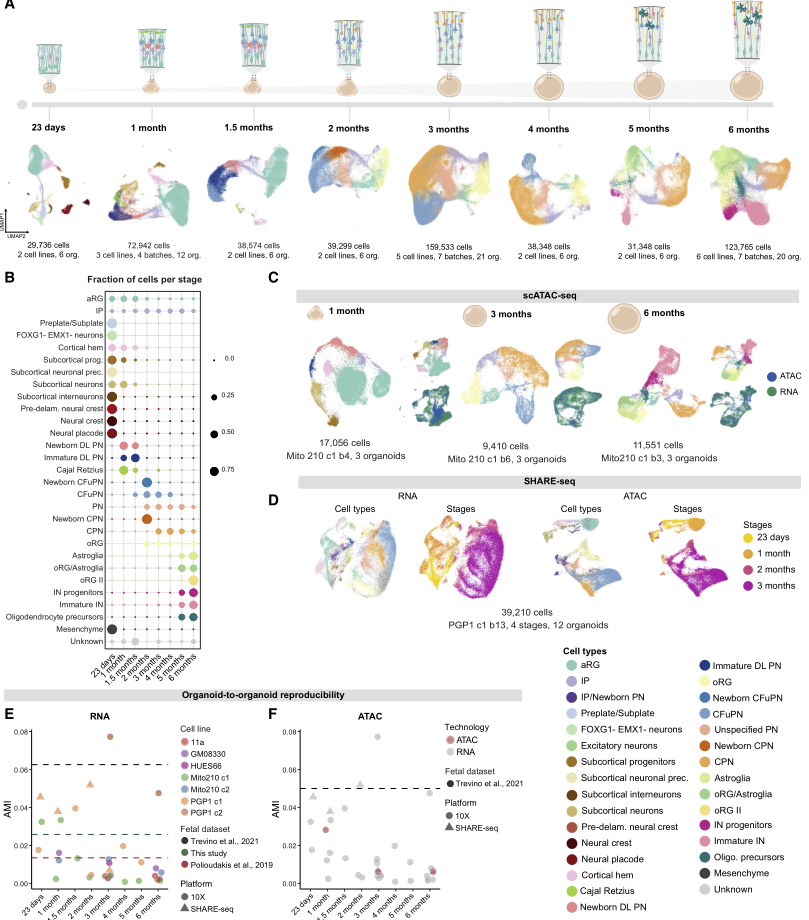
圖1 皮質類器官發育的單細胞轉錄組學和表觀遺傳學景觀
2、類器官發育過程中細胞類型的空間組織
為在其原始空間背景下分析分子和細胞事件,在1個月、2個月和3個月時使用Slide-seqV2進行空間轉錄組分析(圖2)。1個月時,頂端放射狀神經膠質(aRG)祖細胞簇占據類器官的中央和周邊區域(圖2A-2E),Cajal Retzius細胞、中間祖細胞(IP)、新生深層(DL)投射神經元(PN)和未成熟的DLPN相對于aRG位于表層。有趣的是,皮質邊緣區和皮質下區的細胞在很大程度上沒有與皮質細胞類型混合。2個月時,aRG在主要由“非特指PN”群體組成的核心周圍形成了一圈玫瑰花結樣結構(圖2F-2H)。其余的皮質細胞類型主要位于aRG的淺表。外放射狀膠質細胞(oRG)相對于aRG進行了表面定位(圖2H),反映了這些細胞在內源性組織中的相對位置。到3個月時,可識別的結構如玫瑰花結丟失(圖2I-2K),與文獻中研究結果一致。類器官的內核主要由“非特指PN”和aRG構成(圖2K)。這些數據顯示了皮質類器官中細胞類型的動態變化和優先定位,反映了體內觀察到的情況。
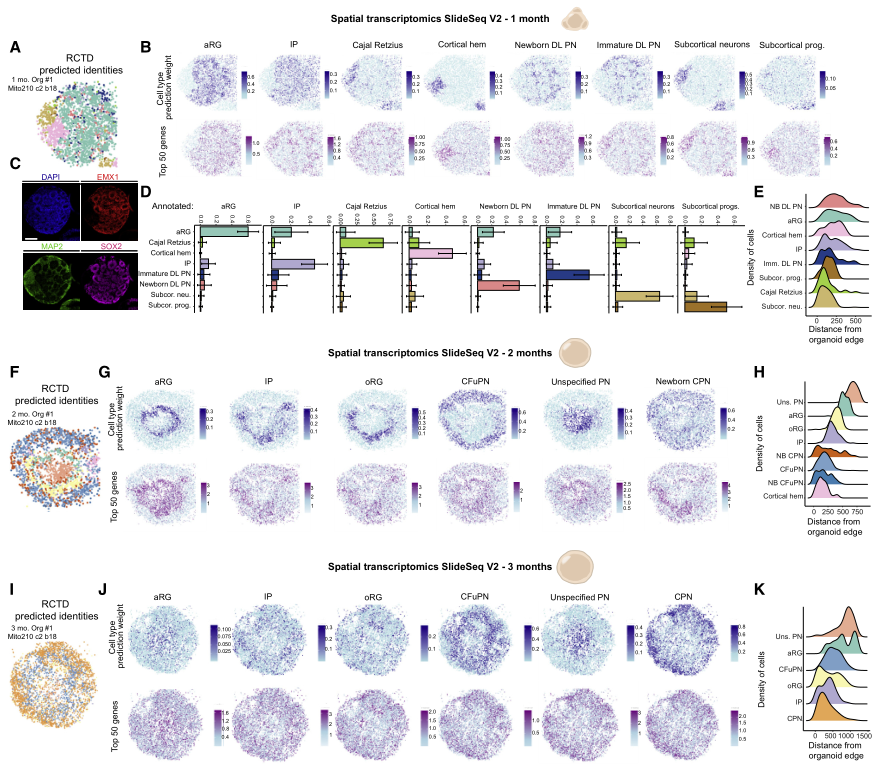
圖2 發育的皮質類器官的空間轉錄組景觀
3、皮質類器官捕獲具有細胞類型和時間特異性的細胞身份獲得的縱向胎兒程序
先前的研究表明皮質器官組織和內源性人類胎兒組織有相似之處,這些相似性是否擴展到所有細胞類型和發育的所有步驟還有待探索。為解決這一問題,作者首先確定了每個階段所有皮質細胞類型的基因表達標簽(圖3A-3B)。接下來,通過在每種細胞類型的可及染色質區域鑒定轉錄因子(TF)基序富集,并將其與同源TF的表達進行比較,研究了假定的細胞類型特異性調節機制。結果證實了已知的發育階段和細胞類型合適的TF及其基序是與相關的細胞群相關(圖3C)。
接下來,試圖確定類器官中識別皮質細胞類型的分子信號與內源性胎兒皮質中定義皮質細胞類型的分子信號有多密切。將類器官數據與兩個已發表的scRNA-seq數據集中的91,844個圖譜和60806個單核RNA測序集進行了整合(圖3D)。以兩種方式評估相似性,這兩種方式都表明類器官和胎兒皮質細胞類型之間高度一致。
首先,應用了基于隨機森林分類器的方法。每個階段的類器官細胞主要由在胎兒細胞上訓練的分類器分配到相應的內源性細胞類型,反之亦然(圖3E-3F)。在用類器官細胞訓練的分類器中,很少有胎兒細胞被分配給aRG和“未指定的PN”類器官細胞(圖3F),這表明這兩個群體與內源性細胞類型的關系不太密切。
其次,使用秩-秩超幾何重疊的方法對類器官和胎兒細胞中每種皮質細胞類型分別定義的分子特征進行了兩兩比較(圖3G)。該方法還顯示了類器官和相應內源性胎兒細胞類型之間的高一致性表達特征(圖3H-3K)。
重要的是,作者發現類器官“未指定PN”的分子標簽,雖然與Polioudakis等人(2019)的胎兒遷移ExNs、Trevino等人(2021)的GluN 1和5簇以及本文胎兒數據集的CPN最接近,但總體上顯示出相對較弱的一致性,主要是由共同的下調基因驅動(圖3J)。類似地,類器官aRG的標記顯示,從體外2個月開始的任何時間點,上調基因與任何胎兒標記都沒有大量重疊(圖3K)。因此,盡管這兩種細胞類型顯示出與內源性細胞的一些總體轉錄相似(圖3E),但它們的轉錄特征與這一胎齡范圍內細胞類型特異性胎兒特征的匹配性相對較弱。
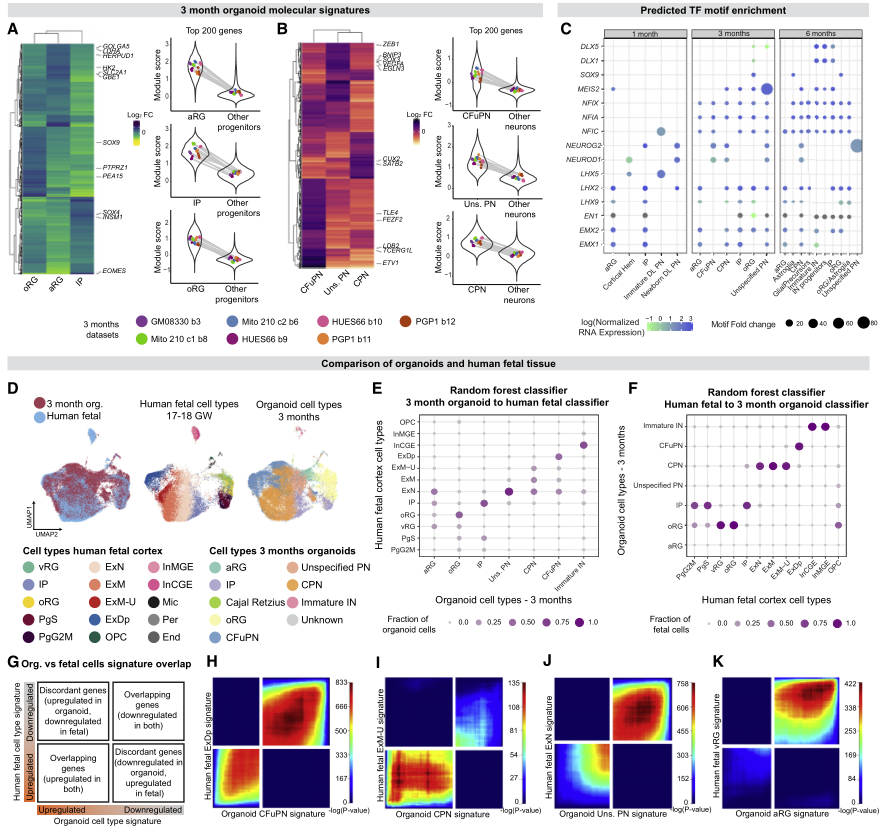
圖3 在具有細胞類型特異性的皮質類器官中建立細胞身份獲取的縱向胎兒程序
4、人類皮質類器官細胞的類型在很大程度上不受代謝狀態的影響
WGCNA確定了一個模塊,其中包括從體外1.5個月開始的糖酵解相關基因,從體外2個月開始,該模塊在與胎兒細胞最不相似的兩個細胞群中特異性富集,即aRG和“未指定PN”(圖4A和4B)。在類器官發育的多個階段,“未指定的PN”和aRG以及其他類型的祖細胞都富集了糖酵解和缺氧基因集(圖4C和4D)。然而,這與在內源性人類胎兒祖細胞子集中觀察到的這些基因集的富集是相似的(圖4E)。因此,在正常皮質祖細胞發育過程中觀察到的代謝狀態也在類器官中相應的細胞類型中觀察到。
對器官組織與內源性胎兒細胞在代謝基因表達方面的差異程度進行的更廣泛研究顯示,所有細胞類型之間的差異都很有限。作者采用三種互補方法: Compass,該方法利用通量平衡分析從 scRNA-seq 數據中模擬單細胞的代謝狀態;RRHO2,識別富集在類器官中上調而在胎兒細胞中下調的基因的過程;以及 MSigDB 代謝基因組的差異表達。在所有三項分析中,類器官和胎兒細胞顯示出相似的結果,只有少數代謝通路,特別是糖酵解和氧化磷酸化,在類器官中富集(圖4F-4G)。值得注意的是,Compass顯示類器官和內源性胎兒細胞很好地聚集在一起,不能根據它們的代謝通量進行區分(圖4F-4G)。
代謝基因在aRG和“非特指PN”群體中的富集可能會干擾類器官細胞類型與其內源性對應的排列,正如文獻里其他皮質類器官模型所建議的那樣。因此,作者從用于訓練隨機森林細胞類型分類器的可變基因中系統地刪除了38個MSigDB代謝基因中的每一個,并評估了對結果細胞分類的影響。對于絕大多數基因集,去除這些基因并沒有顯著改變分配給每種細胞類型的細胞數量(圖4H-4J)。只有兩類代謝過程影響分類:在用3個月的類器官細胞訓練的分類器中,去除與葡萄糖代謝和缺氧相關的基因集顯著增加了分配到“未指定PN”和aRG細胞類型的胎兒細胞數量(圖4H-4J)。沒有其他細胞類型顯示分配增加。重要的是,與凋亡和氧化磷酸化等其他基本亞細胞過程相關的基因未顯著改變任何胎兒細胞類型向類器官細胞類型的分配(圖4H)。

圖4 人類皮質類器官的細胞類型在很大程度上不受代謝狀態的影響
然后,作者利用空間轉錄組學發育圖譜來研究皮質類器官細胞的地形位置和代謝狀態之間是否存在關聯。2個月和3個月的類器官內部區域主要由aRG和非特指PN組成(圖2F-2K,圖5A),這兩個群體的代謝狀態影響了細胞身份的分配。值得注意的是,aRG在1個月和2個月之間變得更加集中,這與我們的發現一致,即aRG的身份在1.5個月開始受到影響(圖5B)。此外,雖然38個MSigDB代謝通路基因集的表達在類器官的直徑中是恒定的,但與缺氧和糖酵解相關的通路在類器官中心的細胞中富集(圖5C-5D)。值得注意的是,位于類器官內部區域的細胞顯示出更高的糖酵解和缺氧基因的表達,這與細胞的特性無關(圖5E)。總之,分析表明,在目前的類器官模型中產生的絕大多數皮質細胞類型的鑒定不受不同的代謝狀態的影響,這只影響了兩種細胞類型(aRG和“非特指PN”)的鑒定,在2-6個月時僅占類器官細胞的3% - 15%。
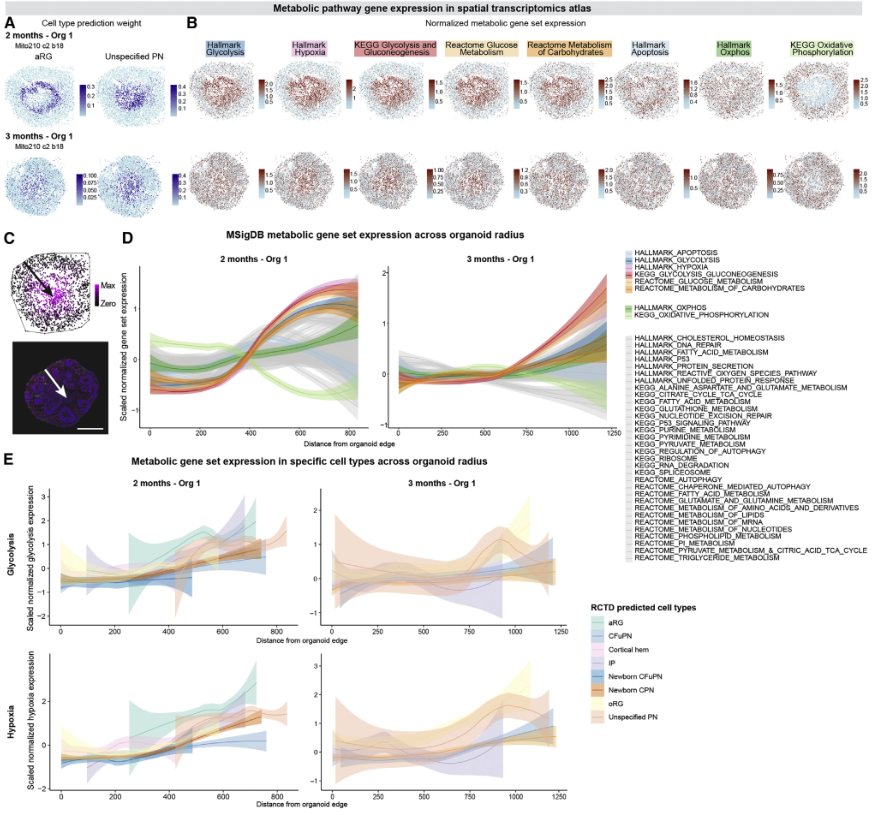
圖5 代謝受損的細胞存在于人類皮質類器官的一個有限的中心區域
5、人類個體皮質細胞類型發育的分子邏輯
前文的結果表明,可以在類器官中建立適當的細胞多樣性,并且細胞身份在很大程度上不受代謝狀態的影響。因此,作者試圖應用這種類器官發育的分子圖譜來理解人類皮質中細胞譜系的建立,以及與譜系決定相關的轉錄事件。作者利用459,711個皮層細胞(不包括未指定的PN)推斷出發展軌跡。為了突出不同時間細胞類型之間的關系,通過轉錄相似性連接了來自相同和連續時間點的細胞簇,同時保留了細胞的已知時間階段(圖6A)。有趣的是,在較晚的時間點,oRG同時連接到神經元和膠質祖細胞(圖6A)。作為一種補充方法,作者應用軌跡推斷算法URD來生成基于偽時間有序細胞轉錄相似性的分支軌跡樹(圖6B)。將根定義為存在于體外23天的最早階段的aRG,而尖端定義為每個皮質譜系的終末細胞類型。在樹中建立的順序反映了細胞的階段和分化狀態,基礎祖細胞在aRG之后,然后分化為神經元(CFuPN,CPN)和膠質(星形膠質細胞,少突膠質細胞前體細胞)譜系。
使用該樹繪制了發育過程中的動態表達變化(圖6C-6D),并確定了與人類皮質神經元亞型相關的譜系特異性基因。為此,將這一人類皮質樹與已發表的小鼠發育皮質URD樹進行了比較(圖6E)。這一分析揭示了人類類器官樹中的譜系特異性基因,而這些基因在小鼠樹中表現出不同的表達模式(圖6F)。例如,SORCS1在人類類器官樹中與CFuPN譜系相關,但在小鼠的多個皮質投射神經元中表達。同樣,PIK3R1在人類中與CPN譜系相關,但在小鼠中表達更廣泛。
為提取與人類細胞命運決定相關的分子程序,確定了與人類類器官URD樹中每個譜系分叉相關的top基因和TF(圖6D)。為了區分控制這些分叉的哺乳動物和人類特有的共享程序,比較了小鼠中與同源分叉相關的TF。值得注意的是,一些被預測在人類神經元命運中發揮作用的候選人類調節因子沒有1:1的小鼠同源物,包括ZNF26和ZNF37A(圖6G)。

圖6 人類皮層細胞類型的分子發育
6、人類胼胝體投射神經元多樣性在胚胎發育過程中出現
作者檢查了樹的終末神經元細胞類型,以研究在成年期CPN基因表達的異質性是否在開發的早期階段就已經可以檢測到。5種不同CPN類型的標記基因在胎兒和類器官CPN的發育早期都有表達,CPN中高表達的標志物也在類器官中高表達,反之亦然(圖7A)。為評估成年期發現的分子多樣性是否也可在早期CPN中觀察到,在成年CPN中確定了5個共變基因程序(模塊),大致代表每種成年CPN類型(圖7B和7C),并在胎兒和類器官CPN中對它們進行了評分。這5個模塊在胎兒和類器官CPN中均有表達。雖然它們的表達并沒有像在成年期中那樣將發育中的CPN細分為5個亞群,但它們開始將早期發育的CPN細分為5個亞群,其中模塊5表現出最獨特的模式(圖7D和7E)。因此,雖然成人CPN的完整表達多樣性還沒有出現,但數據表明CPN的分化是一個開始于早期發展階段的過程。
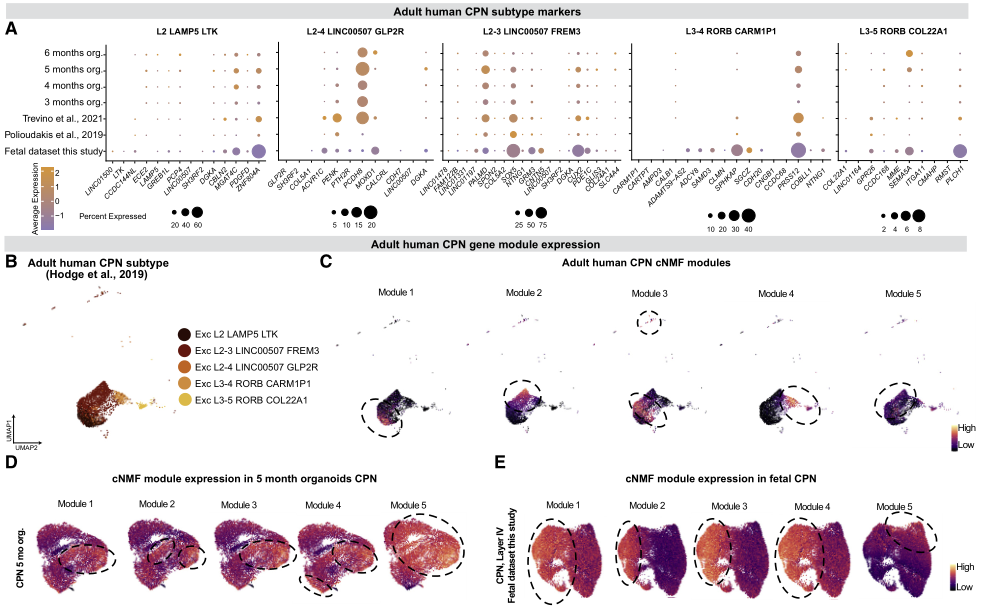
圖7
實驗方法
多能干細胞培養,類器官分化,人胚胎腦組織分離,免疫組織化學,低氧探針,免疫熒光,腦類器官分離,scRNA-seq及其生物信息學分析,單細胞ATAC-seq及其生物信息學分析,SHARE-seq和bulk ATAC-seq及其生物信息學分析,Slide-seqV2及其生物信息學分析,單核RNA測序及其生物信息學分析,大腦皮層類器官和人胚胎的細胞類型比較和匹配分析,WGCNA分析,代謝活動評價,皮質類器官和全腦類器官的比較,代謝基因對細胞分類的影響分析,與代謝相關的表觀遺傳變化分析,軌跡分析,人類胼胝體投射神經元多樣性分析,量化和統計分析
參考文獻
Uzquiano A, Kedaigle AJ, Pigoni M, Paulsen B, Adiconis X, Kim K, Faits T, Nagaraja S, Antón-Bola?os N, Gerhardinger C, Tucewicz A, Murray E, Jin X, Buenrostro J, Chen F, Velasco S, Regev A, Levin JZ, Arlotta P. Proper acquisition of cell class identity in organoids allows definition of fate specification programs of the human cerebral cortex. Cell. 2022 Sep 29;185(20):3770-3788.e27. doi: 10.1016/j.cell.2022.09.010. PMID: 36179669; PMCID: PMC9990683.