






快看過來 這里有鐵死亡的研究新思路
鐵失調已被牽涉到多種神經(jīng)退行性疾病,包括帕金森病(PD)。受影響的大腦區(qū)域經(jīng)常發(fā)現(xiàn)鐵負荷的小膠質細胞,但鐵的積累如何影響小膠質細胞的生理功能并促進神經(jīng)退行性疾病的發(fā)生,目前還不太清楚。在這里,作者表明在一個三聯(lián)培養(yǎng)系統(tǒng)中生長的人類iPSC衍生的小膠質細胞對鐵有高度的反應,并容易受到鐵死亡的影響。此外,鐵過載導致小膠質細胞轉錄狀態(tài)的明顯轉變,與PD死后大腦小膠質細胞中發(fā)現(xiàn)的轉錄組特征相重疊。這種小膠質細胞的反應有助于神經(jīng)退行性改變,因為將小膠質細胞從三聯(lián)培養(yǎng)系統(tǒng)中移除極大延遲了鐵誘導的神經(jīng)毒性。為闡明調節(jié)小膠質細胞中鐵反應的機制,進行了全基因組CRISPR篩選,并確定了鐵死亡的新調節(jié)因子,包括囊泡販運基因SEC24B。這些數(shù)據(jù)表明,小膠質細胞鐵超載和鐵攝取作用在神經(jīng)退行性中起著關鍵作用。本文于2023年1月發(fā)表在《Nature Neuroscience》IF:28.771期刊。
技術路線

主要實驗結果
1、人iPSC來源的三聯(lián)培養(yǎng)物對鐵死亡敏感
為更好地了解鐵信號傳導和鐵死亡在腦中的作用,作者建立了一個由人類iPSC衍生的神經(jīng)元、星形膠質細胞和小膠質細胞的三聯(lián)培養(yǎng)體系(Tri-culture),如圖1A所示。在2周內(nèi),所有的細胞形成了一個復合網(wǎng)絡(圖1B)。為研究鐵過載和鐵死亡的作用,用鐵和RSL3(鐵+RSL3)處理培養(yǎng)物,RSL3是GPX4的抑制劑和已知的鐵死亡誘導劑。通過細胞死亡染料Draq7檢測,雖然單獨使用鐵或RSL3導致的細胞死亡很少,但抑制GPX4和外源性鐵在處理20小時后誘導了大量的細胞死亡,這表明該培養(yǎng)系統(tǒng)可以發(fā)生鐵死亡(圖1C-1D)。鐵+RSL3誘導的細胞死亡可被兩種抗氧化劑鐵死亡抑制劑lip-1和benzothiazine抑制至鐵單獨處理發(fā)水平,這支持鐵死亡是死亡機制(圖1C-1D)。在處理后18小時,有明顯的小膠質細胞死亡,但神經(jīng)元死亡多發(fā)生在20小時左右(圖1C)。星形膠質細胞在任何時間點均未觀察到鐵死亡,這可能與抗鐵死亡的NF2-Hippo通路相關基因的高表達有關。這些結果表明,人三聯(lián)培養(yǎng)系統(tǒng)可作為與神經(jīng)退行性疾病相關的細胞類型的鐵反應和鐵死亡的模型。
為更好地闡明不同類型的細胞是如何對鐵誘導作出反應的,通過scRNA-seq測量了三聯(lián)培養(yǎng)系統(tǒng)中細胞特定的轉錄組變化。分析了刺激后6小時的細胞,這個時間點在鐵+RSL3條件下的明顯死亡誘導之前(圖1c)。在制備細胞的過程中,使用了轉錄抑制劑來保持細胞狀態(tài)。總共對109,100個細胞進行了測序,每個細胞的中位深度為~40,000個讀數(shù)。在五種條件下確定了17個聚類(圖1f)。利用集群基因表達特征和每種細胞類型的幾個已知標記,能夠在所有條件下區(qū)分小膠質細胞、神經(jīng)元和星形細胞(圖1e-g)。進行pseudo-bulk分析,與對照相比,在鐵+RSL3條件下確定了一個鐵蛋白誘導的特征(圖1h)。然后用UCell21檢測了所有17個亞群的鐵誘導特征的富集情況。發(fā)現(xiàn),小膠質細胞亞群2的每個細胞的特征富集程度最大(圖1i)。KEGG途徑分析確定鐵死亡和谷胱甘肽代謝是受影響最大的途徑。此外,幾個免疫相關和骨髓相關的途徑失調,包括趨化因子信號和吞噬體(圖1j)。這些結果表明,與神經(jīng)元和星形膠質細胞相比,小膠質細胞對鐵過載最敏感,可能是鐵中毒級聯(lián)中的一個關鍵角色。
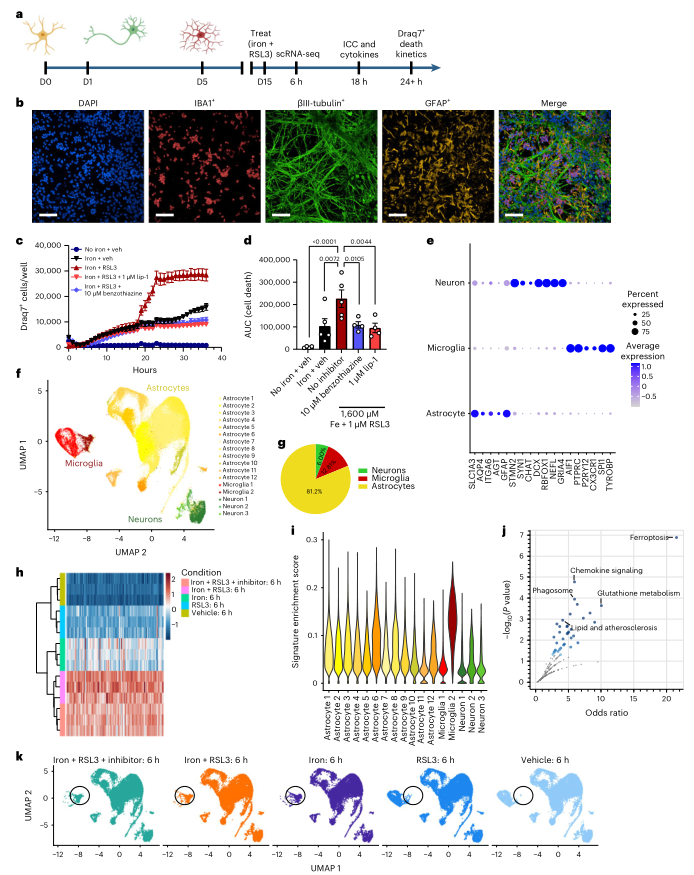
圖1在iPSC三聯(lián)培養(yǎng)中,誘導鐵死亡可引起獨特的轉錄反應和細胞死亡
2、鐵過載和鐵死亡改變小膠質細胞的狀態(tài)
為了解鐵刺激對小膠質細胞狀態(tài)的直接影響,對小膠質細胞進行了進一步分析,發(fā)現(xiàn)了三個獨特的亞群(圖2a-c)。通過檢查富集的基因和使用KEGG途徑分析,確定第2群是一個獨特的鐵死亡相關轉錄組標簽(FAS)亞群。該簇中的細胞富集了鐵死亡和脂質途徑以及p53信號、自噬和內(nèi)質網(wǎng)(ER)壓力(圖2d,e)。FAS亞群在純鐵、鐵+RSL3、鐵+RSL3+benzothiazine的條件下高度富集,并具有pseudo-bulk特征有最高的富集分數(shù)(圖1i,k)。FAS小膠質細胞在鐵+RSL3+benzothiazine條件下持續(xù)存在,表明這種抗氧化劑抑制劑作用于鐵的下游,盡管在很大程度上阻止了鐵死亡。 在對照組中富集的另外兩個小膠質細胞亞群包括一個穩(wěn)態(tài)亞群(簇1)和一個增殖亞群(簇3)(圖2a-b)。
盡管小膠質細胞對鐵過載有明顯的反應,但通過計算每種細胞類型中鐵+RSL3和無鐵+對照組的差異表達基因(DEGs)來研究神經(jīng)元和星形細胞的轉錄反應。在FAS小膠質細胞中被上調的幾個與鐵相關的基因在星形膠質細胞和神經(jīng)元中也差異表達,包括GCLC、GCLM、SLC7A11和TFRC。然而,F(xiàn)AS小膠質細胞的top標記CXCL8在其他細胞類型中沒有表達,總體而言,不同細胞類型的DEGs重疊有限(圖2f)。然而,在神經(jīng)元和星形膠質細胞中,鐵死亡是最主要的激活途徑。FAS小膠質細胞中最顯著的DEG之一是FTH1(圖2d)。FTH1在小膠質細胞中上調,但在神經(jīng)元中不變,在星形膠質細胞中下調(圖2g),表明暴露于誘導鐵中毒的刺激后,鐵的存儲有差異。為證實這一點,進行IHC實驗,發(fā)現(xiàn)暴露于鐵+RSL3后,F(xiàn)TH蛋白水平在小膠質細胞中明顯增加,而不是在星形膠質細胞(圖2h,i)。這些發(fā)現(xiàn)表明,小膠質細胞對鐵暴露有最大的反應,并且在三聯(lián)培養(yǎng)中的鐵存儲量最大。
雖然鐵螯合是小膠質細胞反應的關鍵部分,F(xiàn)AS小膠質細胞也上調CXCL8/IL-8 mRNA(圖2D)。為評估鐵過載是否引起細胞因子分泌的變化,在處理后18小時測定細胞培養(yǎng)上清液中的一系列炎性細胞因子。發(fā)現(xiàn),與對照組相比,鐵處理后IL-8增加了6.8倍,但不顯著,鐵+ RSL3條件下分泌的IL-8顯著增加了7.8倍,benzothiazine或lip-1可部分阻斷IL-8的產(chǎn)生(圖2J)。這些結果表明,鐵過載引起小膠質細胞細胞因子產(chǎn)生和釋放的變化,可能參與了疾病的病理環(huán)境。

圖2與星形膠質細胞和神經(jīng)元相比,誘導鐵死亡引起小膠質細胞狀態(tài)的深刻改變
3、小膠質細胞加劇鐵死亡驅動的神經(jīng)變性
神經(jīng)退行性疾病中含鐵小膠質細胞的存在以及鐵過載后小膠質細胞狀態(tài)的轉變提出了它們可能導致神經(jīng)元死亡的可能性。為研究FAS小膠質細胞在神經(jīng)退行性疾病中的作用,在有或沒有小膠質細胞的情況下建立了星形細胞-神經(jīng)元共培養(yǎng)物(圖3a)。值得注意的是,沒有小膠質細胞的培養(yǎng)物具有明顯的延遲和減少的鐵死亡(圖3b, c)。
為直接評估這種差異是否由神經(jīng)元丟失引起,進行了IHC,發(fā)現(xiàn)只有在含有小膠質細胞的共培養(yǎng)中,神經(jīng)元表面積才顯著丟失(圖3d, e)。還發(fā)現(xiàn),只有當小膠質細胞存在時,神經(jīng)元脂質過氧化才會顯著增加(圖3d, f)。值得注意的是,自由基捕獲抗氧化劑lip-1和fer-1以及鐵螯合劑DFO均可挽救神經(jīng)元丟失,并部分挽救神經(jīng)元和小膠質細胞的脂質過氧化(圖3e, f)。神經(jīng)元表面積與脂質過氧化呈負相關(圖3g),處理24小時后的小膠質細胞存活數(shù)量和鐵死亡處理與神經(jīng)元表面積減少呈正相關,與神經(jīng)元脂質過氧化呈負相關,表明小膠質細胞死亡在神經(jīng)毒性中的作用(圖3h,i)。總之,這些結果表明,小膠質細胞表型的改變和小膠質細胞的死亡產(chǎn)生了一個導致神經(jīng)元毒性的環(huán)境。

圖3小膠質細胞使神經(jīng)元對鐵死亡依賴性變性敏感
4、鐵死亡特征在神經(jīng)退行性疾病中富集
目前缺乏明確的生物標志物來確定鐵的積累是否導致患者的鐵死亡。作者探討了是否能從iPSC三聯(lián)培養(yǎng)系統(tǒng)中發(fā)現(xiàn)PD和ALS患者樣本中富集的鐵死亡特征。分析了3PD患者和3名健康對照組的死后putamen腦組織的snRNA-seq(圖4a)。無偏聚類確定了小膠質細胞、少突膠質細胞前體細胞(OPCs)、少突膠質細胞、神經(jīng)元、星形膠質細胞和內(nèi)皮細胞在測序樣品中的數(shù)量相似(圖4b)。對FAS小膠質細胞數(shù)據(jù)集進行了單細胞加權基因共表達網(wǎng)絡分析(hdWGCNA),以確定五個與鐵死亡有關的基因模塊(圖4c)。FAS小膠質細胞在模塊3、4和5中顯著富集,而穩(wěn)態(tài)小膠質細胞在模塊2中顯著富集(圖4d)。模塊4富集了與鐵失調、谷胱甘肽轉移酶活性、鐵結合和花生四烯酸結合有關的基因(圖4d-f)。與健康對照相比,與鐵相關的模塊4和模塊3在PD小膠質細胞中顯著富集(圖4g)。此外,許多在FAS小膠質細胞中上調的基因在PD小膠質細胞中也上調,包括CXCL8、FTH1、ATG4C和EDEM1。
數(shù)據(jù)顯示,鐵死亡相關途徑,包括那些與FAS小膠質細胞相關的途徑,在死后的腦組織中是上調的,但是,為了研究這種特征是否可以在活著的病人中檢測到,作者分析了健康人和PD參與者的血液RNA-seq數(shù)據(jù),這些參與者參加了加速帕金森病藥物伙伴關系(AMP PD)、帕金森病進展標志物計劃(PPMI)的隊列研究(n = 1,433; 816名帕金森病患者和617名對照組)和帕金森病生物標志物計劃(PDBP)(n = 1,284; 780名帕金森病患者和504名對照組)(圖4h)。PD病例和健康對照組之間的差異性基因表達分析發(fā)現(xiàn)了明顯上調和下調的鐵死亡相關基因,其中許多在三聯(lián)培養(yǎng)FAS小膠質細胞簽名中發(fā)現(xiàn),包括GCLC、GPX4、HMOX1、SQSTM1和SLC7A11(圖4i)。事實上,鐵死亡和IL-8信號傳導是IPA確定的最高失調路徑之一(圖4j)。這些發(fā)現(xiàn)表明,在來自PD患者的血液樣本中存在一個鐵死亡基因特征,并且鐵失調可能在患者體內(nèi)發(fā)生。此外,這些發(fā)現(xiàn)表明,血液樣本有可能被用作外周生物標志物,用于PD和其他神經(jīng)退行性疾病的鐵死亡患者識別和分層。
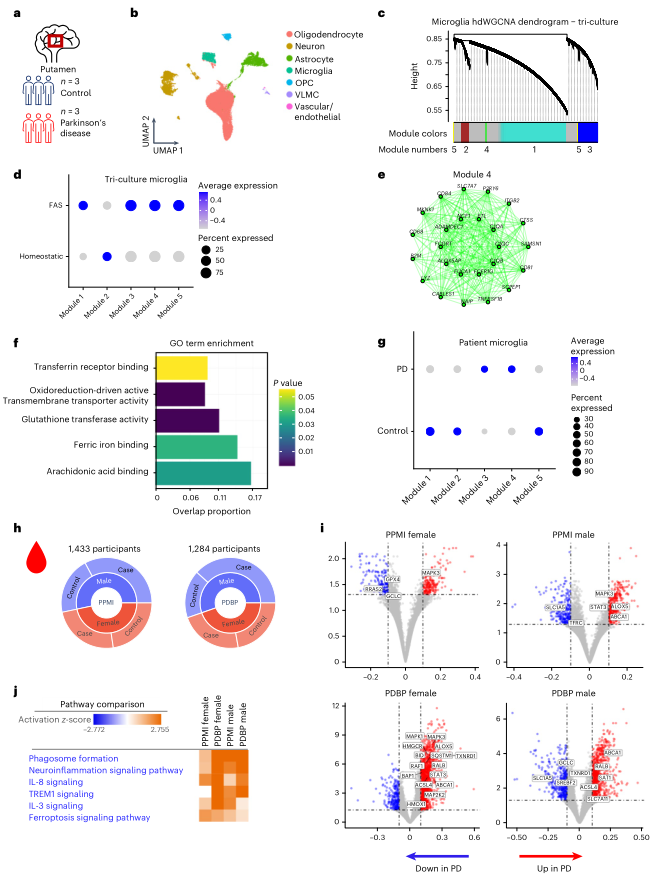
圖4檢測PD患者小膠質細胞的FAS標簽和AMP PD患者血液樣本中的鐵死亡基因標簽
5、CRISPR篩選鑒定小膠質細胞鐵死亡調節(jié)因子
作者使用表達cas9的永生化人類小膠質細胞進行了正向選擇的全基因組CRISPR篩選,以實現(xiàn)全基因組分析所需的可擴展性。使用針對19114個基因的76,612個向導庫,每個基因有四個向導。用兩個單獨生成的病毒庫轉導細胞,用對照或鐵+RSL3處理8小時或24小時,進行較少和較嚴格的選擇(圖5a)。從8小時暴露中發(fā)現(xiàn)了61個命中(圖5b),從24小時暴露中發(fā)現(xiàn)了65個命中(圖5e)。這些數(shù)據(jù)表明,在多種細胞類型中存在一個鐵死亡調節(jié)器的核心網(wǎng)絡,但調節(jié)器可能是細胞類型特定的。
GO分析在兩個時間點都發(fā)現(xiàn)了與ER壓力和運輸以及ER-Golgi囊泡運輸有關的基因(圖5c、d、f、g)。有趣的是,這些途徑在FAS小膠質細胞中也是富集的(圖2d,e)。ACSL4是一個成熟的鐵死亡調節(jié)因子,而SEC24B是本研究中發(fā)現(xiàn)的一個新因子,是唯獨兩個在兩個時間點和庫中都存在的基因(圖5b,e)。SEC24B是COPII介導的從ER到高爾基體的蛋白調節(jié)器。自噬、溶酶體和內(nèi)體途徑也被富集(圖5c、d、f、g)。SEC24B與ER-高爾基體中間室(ERGIC)中的自噬體形成有關,而自噬也與鐵死亡有關通過鐵噬。ER-高爾基體和自噬相關途徑的這種關聯(lián)以及與SEC24B的聯(lián)系促使作者研究SEC24B調節(jié)鐵死亡的潛在機制。
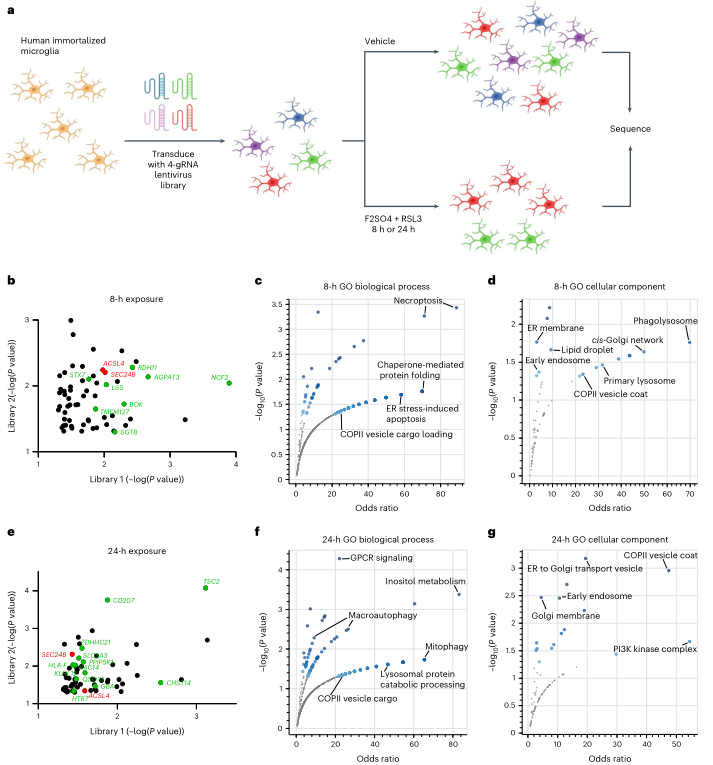
圖5全基因組CRISPR篩選鑒定SEC24B是小膠質細胞鐵死亡的一種新型調節(jié)劑
6、SEC24B敲除可阻止小膠質細胞的鐵死亡
為探索SEC24B作為鐵死亡調節(jié)器的作用,評估了SEC24B基因敲除(KO)的骨髓細胞系對鐵死亡的敏感性。通過qRT-PCR和WB證實了SEC24B缺失,(圖6a,b)。為評估鐵死亡的敏感性,將細胞暴露在鐵+RSL3下,通過Draq7的攝取量測量24小時內(nèi)的細胞死亡。SEC24B KO細胞對鐵死亡有很強的抵抗力,與對照的同源系相比,鐵死亡減少了4倍(圖6c)。SEC24B KO細胞在暴露于鐵中毒刺激2-4小時后,脂質過氧化也明顯減少,脂質過氧化沒有基礎差異(圖(Fig.6d)。這些結果表明,SEC24B是小膠質細胞和巨噬細胞中誘死亡的一個強有力的調節(jié)器。
SEC24B通過結合SEC24A和SEC23B并定位于ERGIC來響應饑餓,從而在自噬流中發(fā)揮作用。一旦形成,該復合體可幫助產(chǎn)生自噬體。有趣的是,SEC24B-COPII定位到ERGIC依賴于PI3K,這是CRISPR篩選中富集的通路(圖5f, g)。為了探索SEC24B和自噬在鐵死亡易感性中的潛在作用,將SEC24B KO HAP1細胞和對照HAP1細胞暴露于自噬抑制劑氯喹,發(fā)現(xiàn)在SEC24B KO細胞中LC3 I/II比值降低,表明自噬流顯著受損(圖6e, f)。為了探索SEC24B是否也調節(jié)鐵自噬,測定自噬條件下的鐵蛋白水平,但發(fā)現(xiàn)鐵蛋白水平?jīng)]有變化(圖6g)。為使用更多與鐵死亡相關刺激,將細胞暴露于erastin,這是一種強烈的鐵死亡誘導劑,已被證明可以驅動鐵平衡的改變和鐵蛋白吞噬。SEC24B KO細胞對erastin誘導的鐵死亡有很強的抵抗力(圖6h),erastin處理導致同源對照組中鐵蛋白和游離易耗鐵明顯增加,但鐵蛋白的增加減少,SEC24B KO細胞中的易耗鐵沒有差異(圖6i,j)。這些發(fā)現(xiàn)表明,雖然SEC24B的缺失導致自噬通量受損,但它很可能沒有通過鐵蛋白自噬來調節(jié)鐵蛋白的攝取。然而,這些結果表明,SEC24B確實改變了易耗鐵池,這是鐵死亡級聯(lián)的一個關鍵上游觸發(fā)因素,這可以解釋SEC24B KO的保護作用。
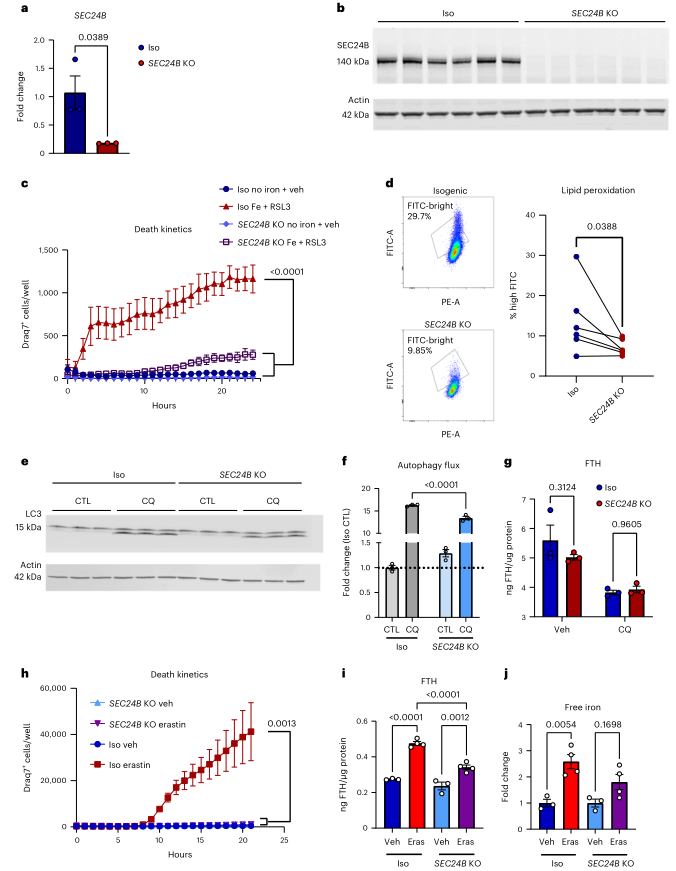
圖6 SEC24B是鐵死亡的有效調節(jié)因子
7、靶向CRISPR的化合物可抑制鐵死亡
鑒于小膠質細胞的鐵死亡在神經(jīng)退行性中的潛在作用,作者試圖找出能抑制小膠質細胞鐵死亡的化合物。為做到這一點,用一個市售的、與鐵死亡有關的庫進行了有針對性的小分子篩選,該庫由546個化合物組成,包括一些已經(jīng)在臨床使用的化合物。將表達Cas9的小膠質細胞系暴露于鐵+RSL3,并與10μM(535/546)或2μM(11/546)的化合物共同處理。與對照相比,任何導致細胞存活率≥70%的化合物都能抑制鐵誘導作用。在546個化合物中,發(fā)現(xiàn)有39個化合物抑制了小膠質細胞的鐵死亡(圖7a-c)。在生物活性數(shù)據(jù)庫ChEMBL57上對這39個化合物進行了分析,以確定它們被預測為作用于哪些途徑。這一分析表明,PI3K信號傳導、GPCR信號傳導和壞死循環(huán)途徑被富集(圖7d)。這些途徑也在CRISPR篩選中被發(fā)現(xiàn)(圖5c,d,f,g)。其他幾個CRISPR目標也與化合物命中的目標有關。例如,兩個細胞色素P450抑制劑(高良姜素和異水楊酸)在化學篩選中被命中,許多細胞色素P450相關基因,包括CYP3A43、CYP46A1和CYP1B1,在CRISPR篩選中被命中。還有多個炎癥相關的基因,如HIF1A、CNR2和NFKBIZ,來源CRISPR篩選的也分別被BAY 87-2243、olivetol和姜黃素及JSH-23命中(圖7e)。因此,藥理篩選確定了一系列的化合物,并驗證了CRISPR篩選中確定的多種途徑和基因。
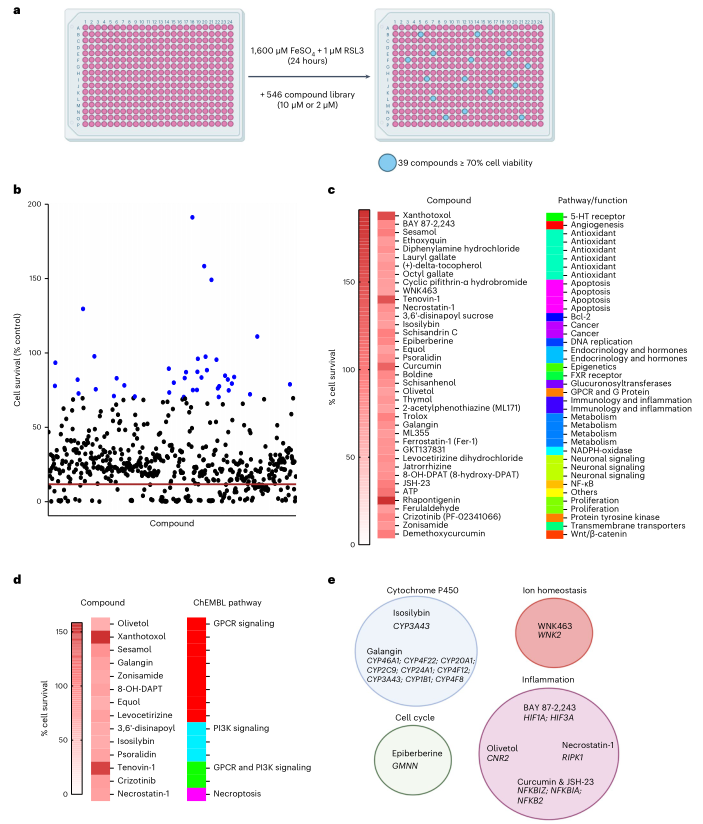
圖7鐵死亡藥物抑制劑的小分子篩選
實驗方法
Tri-culture組裝,Tri-culture鐵死亡處理,單細胞RNA測序(scRNA-seq),Tri-culture的Draq7死亡動力學,永生化小膠質細胞和HAP1細胞系的Draq7死亡動力學,全基因組CRISPR篩選,商業(yè)鐵死亡相關化合物的篩選,
參考文獻
Ryan SK, Zelic M, Han Y, Teeple E, Chen L, Sadeghi M, Shankara S, Guo L, Li C, Pontarelli F, Jensen EH, Comer AL, Kumar D, Zhang M, Gans J, Zhang B, Proto JD, Saleh J, Dodge JC, Savova V, Rajpal D, Ofengeim D, Hammond TR. Microglia ferroptosis is regulated by SEC24B and contributes to neurodegeneration. Nat Neurosci. 2023 Jan;26(1):12-26. doi: 10.1038/s41593-022-01221-3. Epub 2022 Dec 19. PMID: 36536241; PMCID: PMC9829540.