






簡單-直接-分高:SLM2以跨突觸方式塑造突觸特性
突觸特性的確定是神經元環路功能的基礎。“末端選擇”轉錄因子協調協調終端基因電池可以指定細胞類型的特定屬性。此外,泛神經元剪接調節因子被認為與指導神經元分化有關。然而,剪接調節因子如何指導特定的突觸特性的細胞邏輯仍然不清楚。本研究對小鼠海馬中替代剪接調節因子SLM2的細胞類型特異性功能進行了系統的遺傳學分析,證明,SLM2依賴的可變剪接(AS)以跨突觸的方式專門指導突觸特性的規范。本文于20233年2月發表在《Cell Reports》IF: 10.0期刊上。
技術路線:

主要實驗結果:
1、SLM2的結合mRNA編碼突觸蛋白
在小鼠海馬體中,SLM2在谷氨酸能錐體細胞中表達,但也在GABAergic中間神經元亞群中表達。其中包括定向-肺泡腔隙-分子(OLM)細胞CA1,這是一類SST陽性的中間神經元。在神經元細胞核內,SLM2主要集中在核亞結構中(圖1A),讓人聯想起類核小體其由SLM2同源物SAM68組成。然而,在海馬神經元中,只有一小部分亞核結構顯示SLM2-SAM68共定位(圖1B)。為確定SLM2相關的RNAs,在小鼠全腦和海馬樣本上使用了增強型交聯和免疫沉淀(eCLIP)。77%的結合事件發生在內含子,而只有2%映射到外顯子(圖1C)。交叉連接誘導截斷位點(CITS)分析確定了確切的蛋白質-RNA交叉連接位點,這些位點富含UWAA元件(W=U/A;圖1D),這是SLM2在體外識別的motif。使用mCross(一種模擬RBP結合序列特異性和交聯位點的計算方法)進行新的motif識別,顯示UUWAAAA 7-mer是體內被SLM2結合的主要RNAmotif(圖1E)。使用CLIPper和IDR鑒定了復制體中的高置信度SLM2結合事件(圖1F)。對SLM2結合的mRNAs的GO分析顯示,編碼谷氨酸突觸成分的mRNAs有很強的富集性(圖1G)。在全腦樣本的424個高置信度的SLM2目標mRNA中,有110個在SynGO45中被注釋為編碼突觸蛋白,其中有59個突觸前和49個突觸后成分(圖1H)。這些包括泛神經元表達的mRNA,如Nrxn1,2,3、Nlgn1、Lrrtm4、Dlgap1,2、Tenm2和Cadm1,以及優先在GABAergic中間神經元表達的突觸后蛋白,如Erbb4和Gria4。在input樣本中沒有觀察到明顯的峰值,而目標mRNA中密集的UWAAmotif簇與SLM2的結合事件緊密相連(圖1I)。這些實驗揭示了一個編碼突觸蛋白的mRNAs陣列,它們在體內被內源性的SLM2結合。
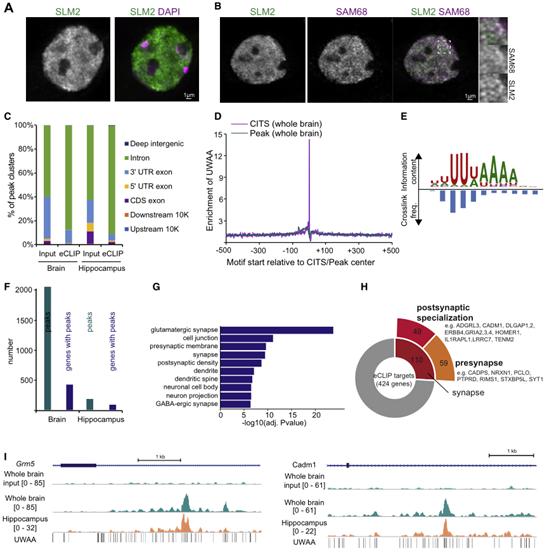
圖1結合SLM2的mRNA編碼突觸蛋白
2、細胞類型特異性SLM2依賴外顯子的鑒定
RBP的作用通常依賴于表達水平。因此,定量了海馬神經元亞群的SLM2免疫反應性(圖2A-B)。SLM2的表達在CA3錐體細胞(Grik4-cre)中表達量最高,在CA1錐體細胞(CamK2-cre)和SST神經元(SST-cre)中明顯表達(圖2B)。超過90%的遺傳標記的CA1和CA3錐體細胞表達SLM2(圖S2C)。相比之下,SLM2在齒狀顆粒細胞中不表達(圖2B)。使用RiboTrap在不同細胞類型中進行條件性消融并繪制各自細胞中的轉錄。分別使用CamK2-cre、Grik4-cre和SST-cre品系在海馬CA1細胞,CA3錐體細胞和SST陽性GABAergic中間神經元中選擇性消融SLM2,分別命名為Slm2△CamK2,Slm2△Grik4,Slm2△SST(圖2C和2D)。SLM2的免疫染色證實了該蛋白在Slm2△SST出生后第16-18天(P16-18)完全喪失,在Slm2△CamK2和Slm2△Grik4在出生后第42-45天(P42-45)75%-90%的cre陽性細胞中完全喪失(圖2D)。
使用RiboTrap親和分離技術,對野生型和敲除型細胞的轉錄組進行了深度測繪。重復樣本之間的差異很小,或者說敲除小鼠與野生型小鼠的RiboTrap樣本之間的差異很小(圖2E),這表明SLM2的損失并不影響這些細胞類型的終端基因電池。散點圖進一步證實在整體基因表達水平上只有很小的改變(圖2F)。最強烈改變的轉錄物是SLM2的類似物SLM1/Khdrbs2,與功能交叉抑制相一致。
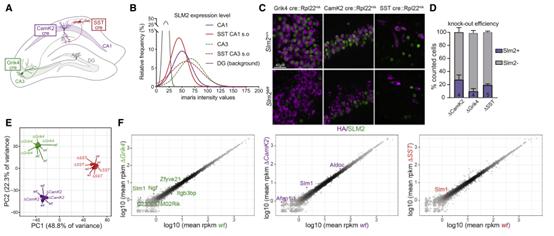
圖2 海馬細胞類型中SLM2的條件消融
SLM2的缺失并沒有廣泛地改變這些細胞類型特異性的剪接特征但改變了少量的AS事件(圖3A),包括Nrxn2選擇性剪接片段4外顯子摻入(AS4)在三個細胞群中增加(圖3A-B)。相比之下,Nrxn3的相應替代外顯子僅在CA3(Grik4)和CA1(CamK2)細胞中被去調控(圖3C)。Syntaxin結合蛋白5-like(Stxbp5l;也叫Tomosyn-2)中相互排斥的替代外顯子e23/e24的去調控是CA1和CA3而不是Slm2△SST細胞中普遍改變的另一個剪接事件(圖3A)。整合eCLIP和RiboTrap剪接分析,發現在去調節的替代外顯子下游500個堿基范圍內有密集的內含SLM2結合事件和UWAAmotif(圖3B和3C)。這表明,SLM2的結合指導了上游替代外顯子的跳躍。
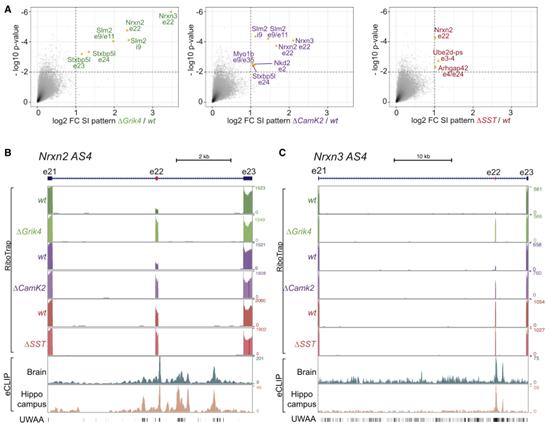
圖3 SLM2直接調控編碼突觸蛋白的mRNA的AS
3、SLM2缺失導致細胞類型特異性突觸表型
考慮到SLM2對結合和調節編碼突觸蛋白的mRNA具有顯著選擇性,所以作者探討了SLM2消融的功能后果。在突觸前CA3錐體細胞中條件消融SLM2導致CA1神經元中Schaffer側支刺激引起的突觸后電流顯著增加(附圖1A-C),因此,從CA3神經元中刪除SLM2足以改變到突觸后CA1錐體細胞的突觸傳遞。從SST中間神經元中條理性敲除SLM2(Slm2△SST)并沒有改變SST陽性中間神經元的內在特性(附圖1C-F),表明SLM2不是這些細胞的特異性所必需的。然而,eCLIP分析發現了大量的SLM2與編碼谷氨酸突觸蛋白的mRNA結合(圖1G-H),所以作者檢測了Slm2△SST細胞的谷氨酸能輸入。結果顯示SST中間神經元的微小興奮性突觸后電流(mEPSC)振幅沒有變化,但觀察到mEPSC頻率顯著升高,表明Slm2△SST細胞上的谷氨酸能突觸密度增加(圖4A-E)。作者調查了對肺泡中假定的CA1軸突進行越來越多的電刺激的AMPAR介導的突觸后反應,發現與較大的突觸數量一致的興奮明顯增加(圖4F)。此外,對同一軸突的40赫茲的刺激導致了短期促進的顯著增加(圖4G)。由于該突觸的短期促進是通過增加遞質釋放介導的,這些結果表明,突觸后SST間神經元的SLM2通過跨突觸機制控制谷氨酸傳導和這些細胞的突觸招募。
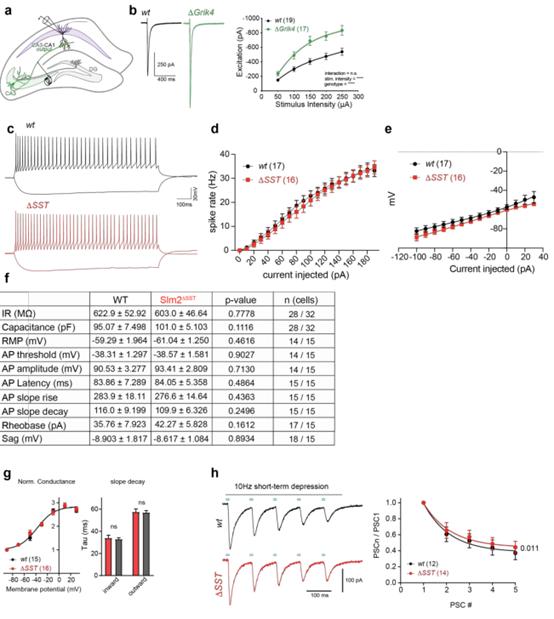
附圖1Slm2條件敲除細胞的電生理分析
通過光遺傳刺激SST中間神經元和全細胞膜片鉗記錄從CA1錐體細胞來進一步分析GABAergic SST中間神經元的輸出突觸。發現Slm2△SST小鼠中光遺傳學誘發的突觸后抑制電流的大小沒有變化(圖4H)。光遺傳學誘發電流的動力學沒有改變,表明突觸后GABA A受體的組裝正常(附圖1G)。應用10 Hz的光遺傳刺激,在野生型細胞中誘導OLM-CA1突觸的抑制,使用該方案,在Slm2△SST小鼠的腦片中觀察到短期抑郁有小幅但顯著的減少(附圖1H)。因此,SST中間神經元的SLM2選擇性缺失導致谷氨酸能驅動增加到OLM中間神經元,與輸出到CA1神經元的突觸基本相似。
研究表明,光遺傳刺激誘導的OLM中間神經元異常激活會損害物體識別實驗中的物體記憶。因此,使用Slm2△SST小鼠進行新的物體識別測試。突變型和野生型小鼠在測試場地的移動能力或與物體交互的總時間方面沒有差異(圖4I)。在測試物體識別記憶時,野生型小鼠探索新物體的時間顯著增加,相比之下,Slm2△SST小鼠與新奇和熟悉的物體互動的時間相似(圖4J-K)。因此,SST中間神經元中SLM2的選擇性缺失與小鼠短期記憶的特定缺陷有關。
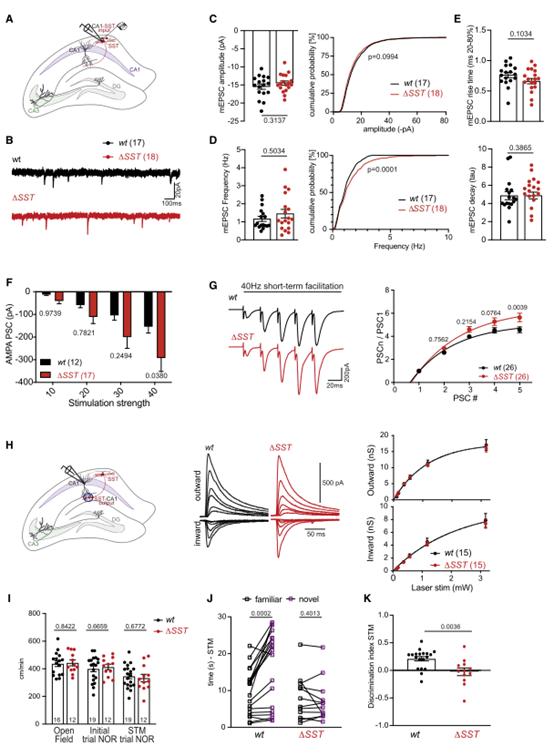
圖4 SLM2通過跨突觸的方式控制突觸的可塑性和功能
總之,本研究表明AS因子SLM2結合編碼突觸蛋白的mRNA;SLM2以細胞類型特異性的方式調節剪接異構體;SLM2缺失導致突觸功能的選擇性非細胞自主改變;在SST中間神經元中缺乏SLM2的小鼠表現出短期記憶缺陷。
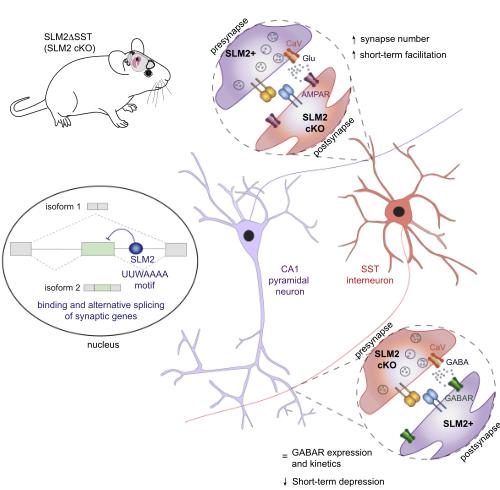
圖形摘要
參考文獻:
Traunmüller L, Schulz J, Ortiz R, Feng H, Furlanis E, Gomez AM, Schreiner D, Bischofberger J, Zhang C, Scheiffele P. A cell-type-specific alternative splicing regulator shapes synapse properties in a trans-synaptic manner. Cell Rep. 2023 Feb 28;42(3):112173. doi: 10.1016/j.celrep.2023.112173. Epub ahead of print. PMID: 36862556.