






成年大鼠睪丸間質細胞去除和修復后的單細胞RNA測序
精子發生是一個高效、復雜和高度組織化的增殖和分化過程,它依賴于多種因素,包括間質細胞產生的睪酮。盡管睪酮在精子發生中所起的關鍵作用已經得到了很好的認識,但其作用機制仍不完全清楚,部分原因是無法特異性和準確地監測發育中的生殖細胞內對睪酮的依賴變化。目前,有研究首次提供了成年大鼠睪丸細胞的單細胞RNA測序數據。該研究發表于《SCIENTIFIC DATA》,IF:8.501。
技術路線:

主要研究結果:
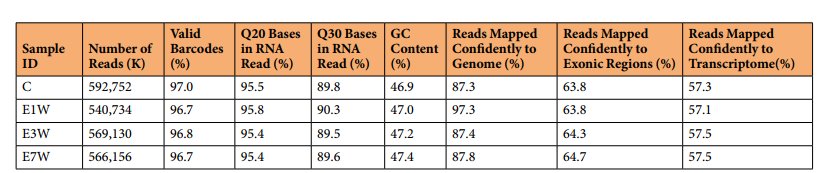
作者使用10x Genomics Chromium平臺構建單細胞RNA-seq文庫,并在Illumina HiSeq PE150平臺進行測序(圖1a)。實驗包括四個睪丸治療組,覆蓋成年大鼠的一個完整的間質細胞耗竭和替代周期。通過血清睪酮測定驗證了乙烷二甲烷磺酸鹽(EDS)誘導的Leydig細胞消除(E1W)和再生(E3W和E7W)的過程(圖1b)。對于測序過程,飽和曲線分析表明,4個處理組的測序深度相當,足以檢測到每個細胞中最多的基因(圖1c,d)。測序的詳細數據質量指標列于表1。所有治療組的總讀長均大于540 M。檢測到的有效條形碼在96.7% ~ 97.0%之間。基因組上的reads在87.3% ~ 97.3%之間,轉錄組上的reads在57.1% ~ 57.5%之間。4種治療的指標非常相似,反映出由于技術原因引入的偏倚很小。
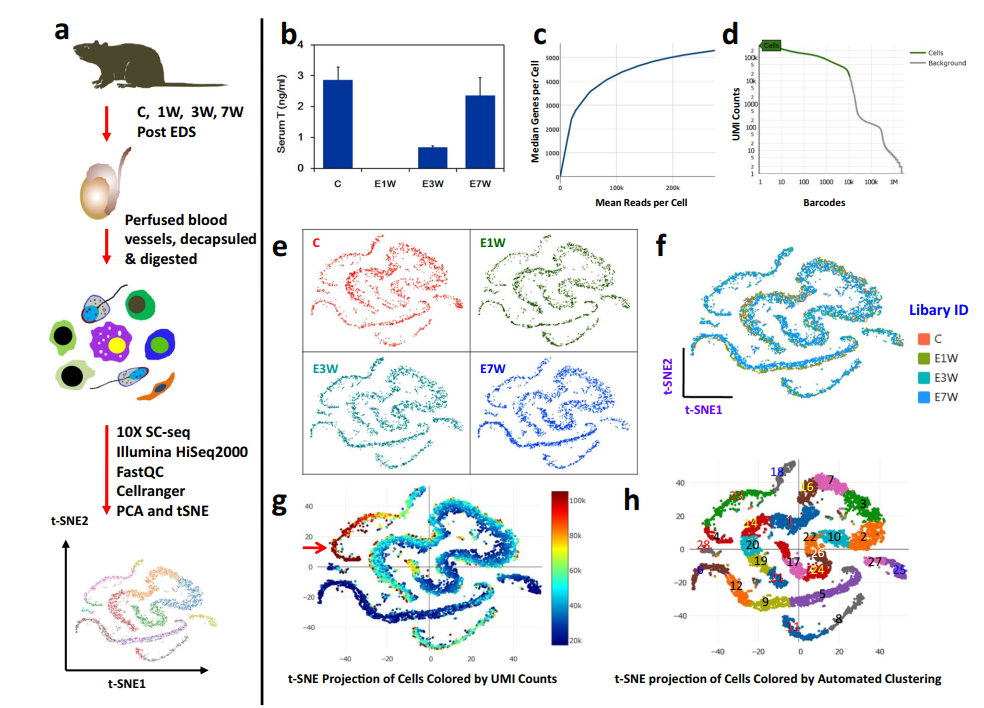
圖1 EDS處理和未處理的大鼠睪丸細胞的scRNA-seq分析
表1 FASTQ文件的詳細QC
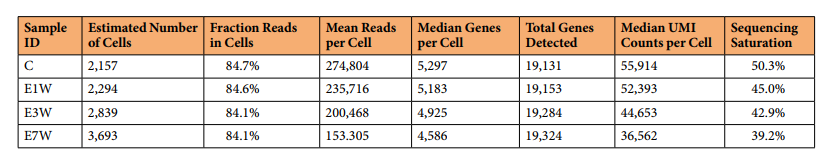
基于細胞的詳細測序統計量列于表2以及圖1d。獲得的細胞數量估計在2,157 ~ 3,693個之間。每個細胞的中位UMI計數在36,562 ~ 55,914之間,每個細胞檢測的中位基因在4,586 ~ 5,297之間。除E7W組捕獲的細胞比其他組多外,其他4個處理組的這些指標非常相似。線粒體轉錄本的百分比是細胞凋亡的粗略指標,而血紅蛋白的百分比則是污染紅細胞的指標。在這個的實驗中,很少有細胞超過5%線粒體轉錄本和0.01% HB轉錄本這一常用閾值,表明純凈和活細胞的質量高。總體而言,除E7W因組內細胞數較多而檢測到的每個細胞的基因中位數略低外,其他指標在4組間非常相似,反映出很少的技術差異。
表2 基于細胞的測序統計
為了進一步比較4個治療組之間的數據一致性,通過PCA將各組的特征維度進行降維,然后通過t-SNE在2個維度上進行投影(圖1f)。令人驚訝的是,各組之間的細胞分布沒有顯著差異(圖1e)。因為睪丸中各種生殖細胞群的RNA含量有很大的差異,在粗線精母細胞中發現的水平最高 (圖1g和2)。包含可檢測基因最多的部分(圖1g,紅色箭頭)正是粗線精母細胞所在的區域(圖2a,c)。通過自動圖形聚類,作者發現了28個亞簇,這表明該數據集在生殖細胞發育階段的識別和表征、與各階段相關的新標志物以及細胞間涉及的特定旁分泌因子方面具有豐富的潛力(圖1h)。

圖2 通過scRNA-seq鑒定的睪丸細胞類型的標志物和屬性概述
為了顯示數據的質量,根據圖形模式和預定義的集群數量對單元格進行了初步的聚類。借助來自大鼠、小鼠和人類的睪丸細胞標志物,作者能夠將28個基于圖的細胞簇合并為包含5個生殖細胞階段的7種主要細胞類型:精原細胞(SPG)、精母細胞(SPC)、圓形精子細胞(RSPT)、伸長精子細胞(ESPT)、濃縮精子細胞(CSPT)和2種體細胞類型Sertoli和Leydig(圖2a)。通過熱圖總結了每種細胞類型的前18個差異表達基因(圖2b),其中5個代表性基因以名稱顯示。集群的一些特定標記也顯示在t-SNE投影(圖2c)或小提琴分布(圖3a)。雖然大多數標記只標記一個特定的發育中的生殖細胞階段或特定的體細胞類型,但其他基因,如眾所周知的生殖細胞標記物Ddx4,標記了除凝集后期以外的整個生殖細胞群。此外,其他基因,如Crisp2或Rps16,在所有類型的睪丸細胞中普遍表達。

圖3 標志基因和睪酮敏感基因的分布
作者又進一步分析了每種細胞類型的知名標記基因的表達模式(圖s7 ~ s8)。Dazl、Sohlh2和Elavl3是已知的SPG標記,在本研究中,這些標記只在聚類中表達(圖S7)。Phf、Id1和Ngfr在SPC中富集,Lrat和Spag在SPT中富集。Tnp和Prm都是眾所周知的標記物。所有這些都被發現在本研究中對應的聚類中特異性表達(圖S7, S8)。另外,兩個主要的體細胞群,Sertoli細胞和Leydig細胞,只表達眾所周知的標記物Clu和Gstm6的支持細胞和Cyp17a1和Cyp11a1的Leydig細胞。所有這些特異的表達標記都支持細胞分類準確的結論。
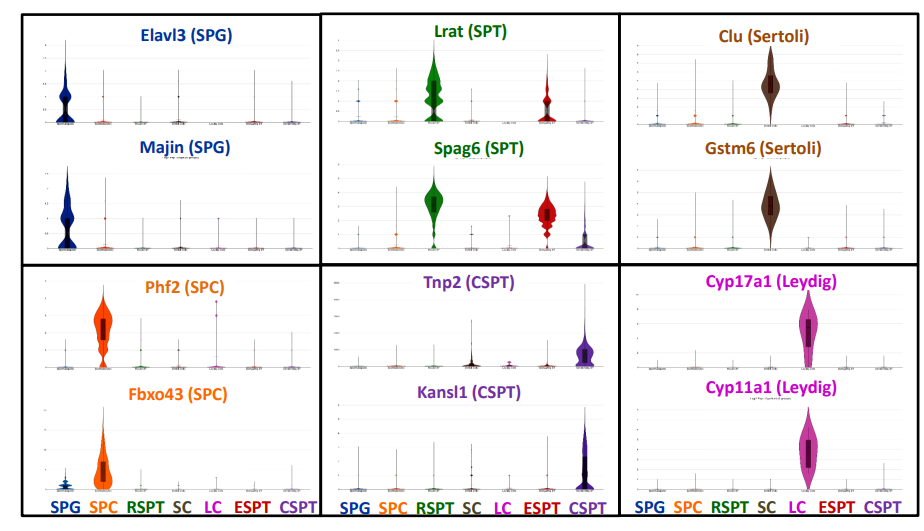
圖S7 不同睪丸細胞特異性表達基因的小提琴圖

圖S8 不同生殖細胞特異性表達基因的小提琴圖
另一個眾所周知的SPG干細胞基因是Gfra1。該基因并不是由SPG表達,而是由SPT高表達(圖S9A, B)。SPT在大鼠中高表達(圖S9C)。
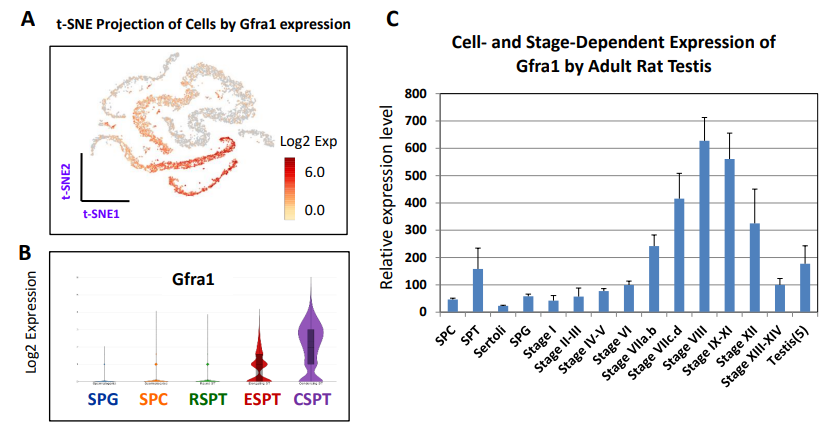
圖S9 Gfra1在不同發育階段生殖細胞中的表達
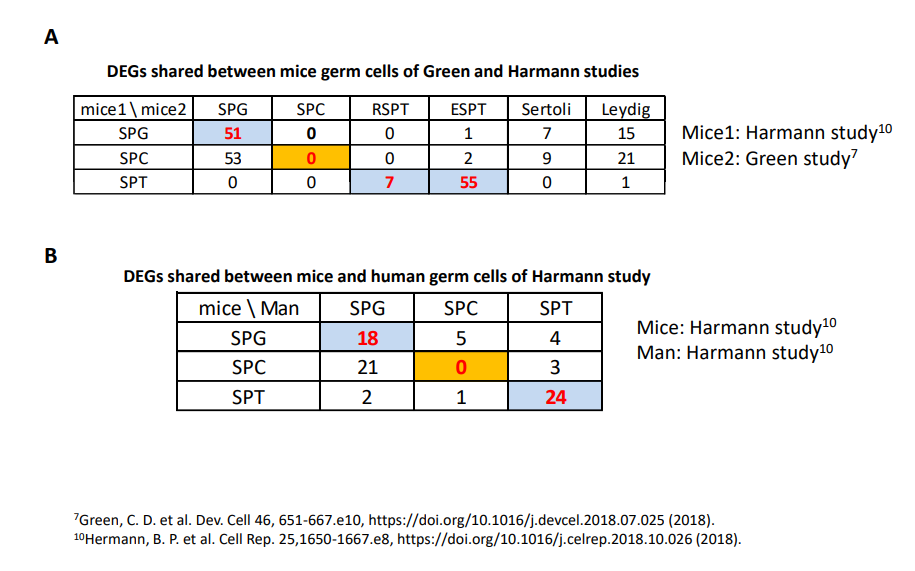
為了進一步檢驗大鼠睪丸細胞的分類在多大程度上與最近對小鼠和人類睪丸細胞的研究一致,作者計算了在本次研究的前200個富集基因和已發表研究的基因列表中每種細胞類型的共享基因的數量。如補充表S1所示,該研究與Green等的研究在每種細胞類型的DEGs數量上是匹配的。體細胞的基因匹配率高于生殖細胞,發育晚期的生殖細胞基因匹配率高于發育早期。不同細胞類型之間共享的基因比相同細胞類型之間共享的基因要少得多。該研究和Hermann的小鼠和人類研究之間的所有細胞也是如此,但小鼠SPC除外(表S1和S2)。這些結果表明,除了小鼠SPC之外,3項研究中其他所有細胞類型都有很高的匹配度,這表明在不同物種的精子發生中有共同的發育和調控機制。
表S1 3項scRNA-Seq研究中睪丸主要細胞的前200個差異表達基因(DEGs)中共享的基因數量
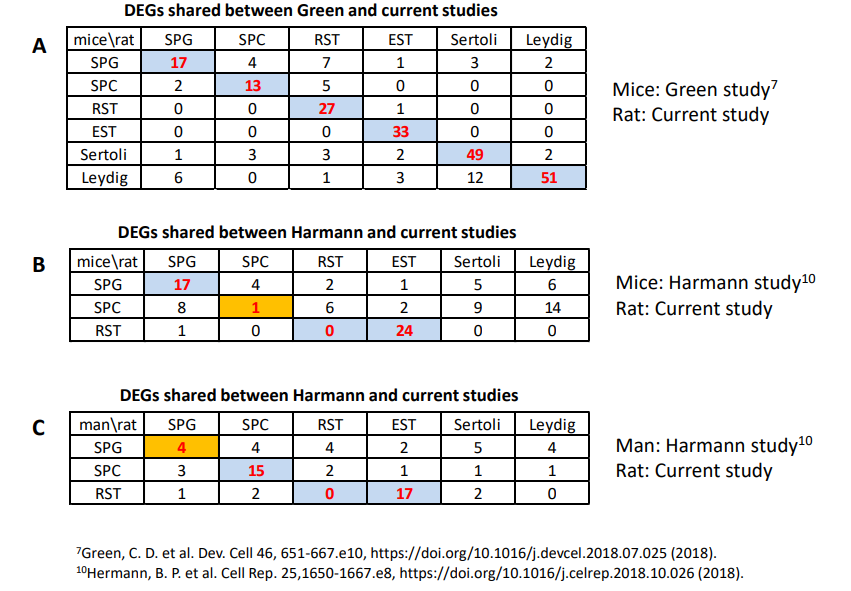
表S2 2項scRNA-Seq研究中睪丸主要細胞的前200個差異表達基因(DEGs)中共享的基因數量
該研究實驗設計的一個顯著特點是睪丸內睪酮的耗竭和恢復。這樣一個動態的周期提供了一個機會來分析睪酮對維持和恢復精子發生的作用。為了檢測睪酮的缺失和恢復是否影響生殖細胞中任何基因的表達,比較4個治療組的基因表達(圖3b)。在1周(E1W)或3周(E3W)睪酮撤退時,Noxred1和Pdia5等基因被抑制(藍框),但在7周(E7W)時恢復正常水平。有趣的是,Noxred1在第1周開始反彈,而Pdia5在第3周開始反彈,這表明可能涉及不同的調控機制。此外,這兩個基因的變化主要是在圓形精子細胞群體中,盡管這兩個基因的表達并不局限于該群體。除了生殖細胞,支持細胞對EDS處理的反應中還有許多基因表達差異(圖3c)。有些基因在EDS治療后1和/或3周被完全抑制,到7周時完全恢復到control (C)水平。此外,在EDS治療后1周和/或3周,一些未被對照組表達的基因被激活。這些基因在EDS治療7周后被關閉。
總結:
這些數據集的主要分析進一步支持了數據的質量,以及探索新的生殖細胞特征、與特定發育階段相關的標志物和細胞-細胞相互作用中的旁分泌因子的潛力,特別是睪酮在精子發生中的作用。
用途:
該數據集可用于(1)在單細胞水平上識別和驗證大鼠睪丸細胞中的新基因和轉錄本,(2)開發更全面的大鼠睪丸單細胞轉錄組注釋系統,(3)識別與大鼠精子發生特定階段相關的新基因調控網絡,以及(4)發現生殖細胞和體細胞中睪酮依賴的基因。