






HMGA2通過腫瘤相關巨噬細胞招募促進結直腸癌進展
腫瘤相關巨噬細胞(TAMs)通常表現為腫瘤前M2樣表型,通過其免疫抑制活性強烈影響結直腸癌(CRC)的進展。HMGA2是一種癌蛋白,在CRC細胞中異常過表達。然而,腫瘤來源的HMGA2調節結直腸癌腫瘤微環境的機制尚不清楚。我們發現,在體外和體內,癌細胞中的HMGA2促進巨噬細胞招募和M2極化。HMGA2直接與STAT3啟動子結合激活其轉錄,進而誘導CCL2分泌,從而促進巨噬細胞募集。腫瘤細胞中HMGA2的表達和間質中CD68的表達之間存在很強的正相關。在所有患者或遠處轉移陰性的亞組中,CD68表達升高的患者的總生存期較差。我們的工作揭示了CRC中HMGA2/STAT3/CCL2軸和巨噬細胞招募之間的聯系。這些發現為靶向CRC中的HMGA2/STAT3/CCL2軸提供了一種新的治療選擇。本文于2022年1月發表于“Theranostics”(IF=11.556)。
技術路線

結果
1)在CRC細胞中敲除HMGA2可抑制TAM浸潤、M2極化和CCL2的分泌
利用CRISPR/Cas9技術,我們用特異性sgRNA在小鼠CRC細胞系(MC38和CT26)中產生了穩定的Hmga2敲除(Hmga2- KO)細胞。Western blot分析驗證了敲除Hmga2的有效性(圖1A)。C57BL/6小鼠皮下接種對照或Hmga2-KO MC38細胞(MC38-NC和MC38-sgA2),BALB/c小鼠皮下接種CT26-NC或CT26-sgA2細胞(圖1B)。結果顯示,sgRNA介導的敲除Hmga2在C57BL/6和BALB/c皮下腫瘤模型中均顯著抑制腫瘤生長(圖1C-D)。
M2巨噬細胞可以促進腫瘤的生長和發展。為了闡明Hmga2在TAM招募和極化中的作用,我們采用流式細胞術量化巨噬細胞(CD11b+F4/80+)和M2巨噬細胞(CD11b+F4/80+CD206+)的百分比。攜帶Hmga2敲除MC38腫瘤的小鼠,浸潤性CD11b+F4/80+巨噬細胞和CD11b+F4/80+CD206+ M2巨噬細胞減少(圖1E),提示Hmga2敲除的抗腫瘤作用可能與CRC中TAMs的招募和極化有關。同樣,在Hmga2敲除CT26腫瘤的小鼠中也觀察到了類似的結果(圖1F)。這些結果表明,Hmga2在體內促進TAM招募和M2極化。
CCL2是一種重要的趨化因子,有助于巨噬細胞招募和浸潤。為了探究CCL2是否受Hmga2調控,我們采用qPCR檢測CCL2的表達,ELISA檢測CCL2的分泌。如圖1G和1I所示,與對照組相比,MC38-sgA2細胞腫瘤中CCL2的表達減少,產生的CCL2減少。同樣,通過qPCR(圖1H)和ELISA檢測(圖1J),在CT26皮下腫瘤模型中,Hmga2的缺失抑制了CCL2的表達和分泌。
接下來,為了探討TME中CRC細胞中Hmga2缺失與巨噬細胞極化之間的關系,我們評估了M1相關(TNF-α和IL-12b)和M2相關(TGF-β)細胞因子的表達。如圖1G-H所示,qPCR結果顯示,CRC細胞敲除Hmga2后,MC38和CT26異種移植瘤中TNF-α和IL-12b的表達均增加,TGF-β水平明顯降低。在MC38和CT26皮下腫瘤模型中,ELISA進一步證實Hmga2-KO腫瘤組織中TNF-α增加和TGF-β分泌減少(圖1I-J)。總的來說,在CRC細胞中敲除Hmga2可以抑制TAM浸潤、M2極化和CCL2的分泌。

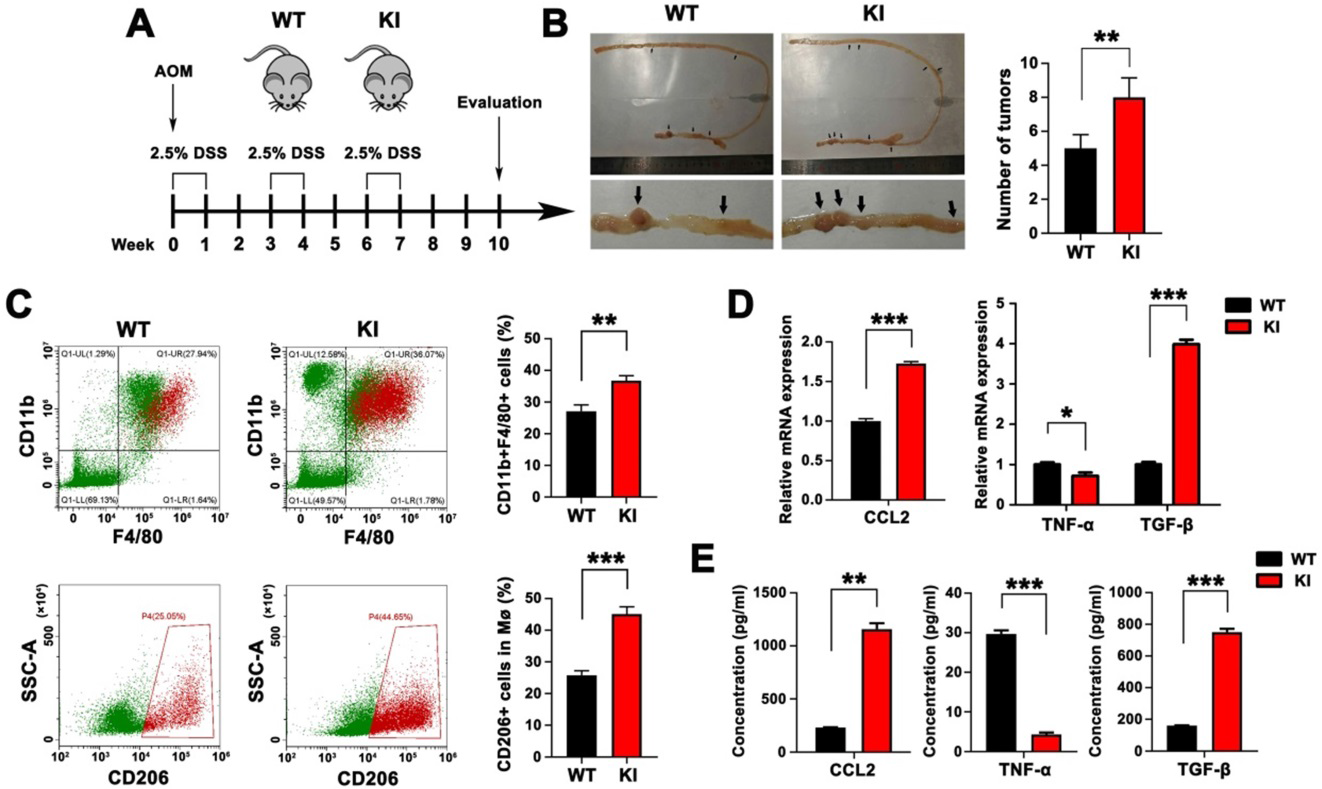
2)在AOM/DSS模型中,HMGA2腸上皮特異性敲入促進TAM浸潤、M2極化和CCL2分泌
我們試圖了解在CRC腫瘤發生過程中,Hmga2的腸上皮特異性敲入(KI)在TAM浸潤和M2極化中的參與。我們使用WT和腸上皮特異性Hmga2 KI小鼠,隨后用AOM和DSS治療誘導結直腸腫瘤(圖2A)。如圖2B所示,Hmga2 KI小鼠比WT小鼠在腸道內發生的腫瘤更多。與WT小鼠相比,在給藥AOM/DSS后,Hmga2 KI小鼠腸道組織中CD11b+F4/80+巨噬細胞和CD11b+F4/80+CD206+ M2巨噬細胞的比例增加(圖2C)。此外,通過qPCR和ELISA檢測介導巨噬細胞浸潤的趨化因子CCL2水平(圖2D-E)。我們發現Hmga2 KI可顯著上調小鼠腸道組織中CCL2的產生。qPCR結果顯示,AOM/DSS處理后,Hmga2 KI小鼠腸道組織中M1細胞因子(TNF-α)的表達顯著降低,M2細胞因子(TGF-β)的表達顯著增加(圖2D)。這些結果經ELISA證實(圖2E)。綜上所述,腸道上皮特異性Hmga2 KI通過促進TAM浸潤、M2極化和CCL2分泌來調節TME。
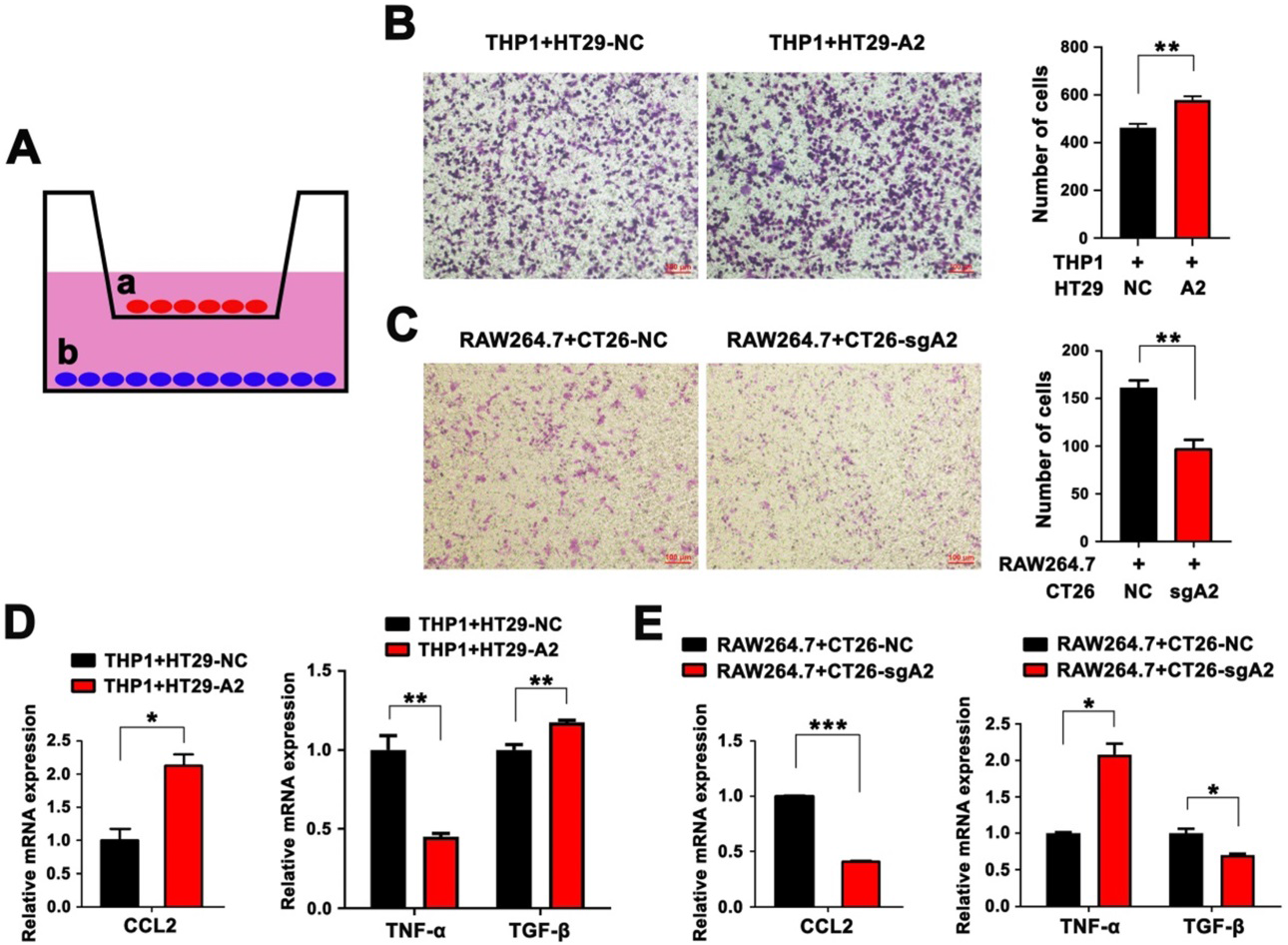
3)HMGA2促進體外巨噬細胞招募、M2極化和CCL2分泌
為了進一步說明HMGA2在體外是否有助于TAM招募,我們利用Transwell共培養系統。如圖3A所示,PMA分化的THP1人單核細胞與HMGA2過表達或不表達的HT29人結直腸癌細胞(HT29-NC和HT29-A2)共培養,RAW264.7細胞與HMGA2敲除或不敲除的CT26小鼠結直腸癌細胞(CT26-NC和CT26-sgA2)。有趣的是,THP1與HT29-A2共培養比HT29-NC細胞表現出更高的遷移能力(圖3B)。相反,與CT26-sgA2細胞共培養時,RAW264.7的遷移能力顯著減弱(圖3C)。隨后,我們通過qPCR檢測CRC細胞中CCL2的表達水平,發現HMGA2過表達會上調HT29細胞中CCL2的表達(圖3D),而HMGA2 KO會下調CT26細胞中CCL2的表達(圖3E)。此外,HT29-A2條件培養基培養的THP1細胞中TNF-α的表達下降,而TGF-β則呈現相反的趨勢(圖3D)。與對照組相比,CT26-sgA2條件培養基處理RAW264.7細胞后,TNF-α水平升高,TGF-β水平降低(圖3E)。這些研究進一步證實HMGA2在體外促進巨噬細胞招募、M2極化和CCL2分泌。
既往研究表明,STAT3在腫瘤免疫耐受中起關鍵作用。為了研究HMGA2是否通過STAT3依賴機制調控TME中TAMs的免疫抑制,我們分析了CRC中HMGA2與STAT3的關系。如圖4A所示,shRNA介導的Hmga2敲低導致MC38和CT26細胞中總和磷酸化的Stat3 (pStat3Tyr705)蛋白水平下降。Western blotting結果顯示,sgRNA介導的敲除Hmga2顯著抑制了MC38和CT26細胞中Stat3和pStat3Tyr705的表達(圖4B)。相反,與對照組相比,Hmga2過表達的LoVo和HT29細胞中STAT3和pSTAT3Tyr705的表達上調(圖4C)。此外,我們將對照或過表達Hmga2的載體轉染到對照或Hmga2缺失的CT26細胞中(CT26-NC、CT26-sgA2、CT26- sgA2 +NC和CT26-sgA2 +A2)。如圖4D所示,Hmga2過表達恢復后,敲除Hmga2后減少的Stat3和pStat3Tyr705的表達量逆轉。為了更好地了解Hmga2和Stat3在體內的調控機制,我們通過Western blotting檢測了在WT和KI小鼠腸道組織中Stat3、pStat3Tyr705和Hmga2的表達。如圖4E所示,我們發現Hmga2的敲入提高了Stat3和pStat3Tyr705的水平。
為了確定HMGA2是否能在轉錄水平激活STAT3,我們進行了熒光素酶和ChIP檢測。如圖4F所示,我們將5個人類STAT3啟動子片段克隆到pGL3載體上,分別為-139/+133、-1555/-140、-850/-140、-1555/-851和-1555/+133。結果顯示,HMGA2過表達顯著刺激了STAT3啟動子區域-1555/-140、-850/-140、-1555/-851和-1555/+133的熒光素酶活性,說明-1555/-140片段可能參與了HMGA2調控STAT3轉錄的過程(圖4F)。為了進一步證實,我們采用ChIP實驗驗證了其直接調控機制,并確定了結合位點的位置。如圖4G所示,HMGA2直接與STAT3啟動子結合,且HMGA2結合位點主要位于-815至-546之間的STAT3啟動子區域。接下來,我們分別突變了STAT3啟動子段側翼的三個Hmga2結合位點(-815/-546)。圖4H顯示的結果表明,轉染含有突變1(-743/-730,從TAATTACTCTATTT到TAGCCACTCTACGT)和突變3(-585/-576,從TATCTAACTA到TCTCGCTA)的結構體后,熒光素酶的誘導活性顯著減弱。這些結果表明,HMGA2通過直接結合STAT3的-743/-730和-585/-576啟動子區域,增強了STAT3的轉錄。

5)CRC細胞中HMGA2過表達增強CCL2表達和巨噬細胞遷移增加依賴于STAT3
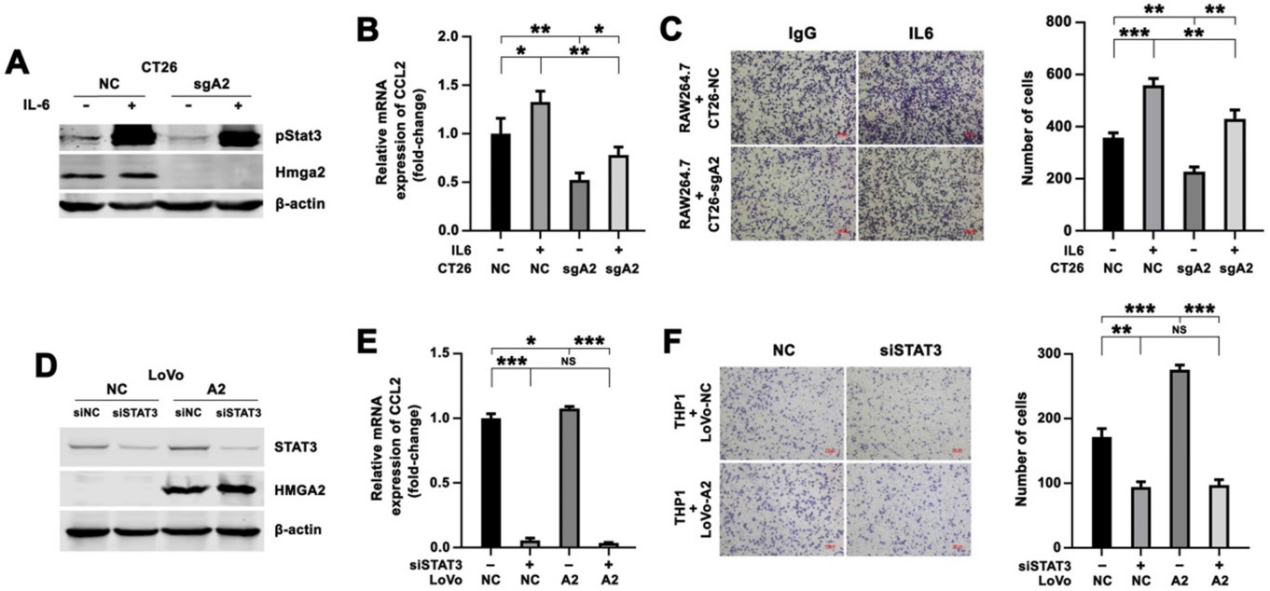
我們通過IL6處理CT26-NC和CT26-sgA2細胞,刺激Stat3,然后與RAW264.7細胞共培養,確定Hmga2是否以Stat3依賴的方式調控CCL2的表達。IL6處理導致pStat3Tyr705表達增加(圖5A), CRC細胞中CCL2表達強烈上調(圖5B), RAW264.7細胞遷移增強(圖5C)。此外,將靶向STAT3的siRNA引入LoVo-NC和LoVo-A2細胞,通過Western blotting評價STAT3抑制效果(圖5D)。我們觀察到,在LoVo-NC細胞中,siSTAT3組CCL2的表達低于對照組,說明STAT3上調了CCL2的表達(圖5E)。我們還發現HMGA2過表達導致CCL2的表達增加,但引入靶向STAT3的siRNA后,CCL2的誘導被取消(圖5E)。在Transwell共培養體系中觀察到一致的結果。LoVo細胞中STAT3沉默導致THP1細胞遷移減少(圖5F)。與LoVo-NC細胞共培養相比,THP1細胞與LoVo-A2細胞共培養顯示出更強的遷移能力,但通過引入STAT3 siRNA,這種增加的遷移被消除(圖5F)。這些結果表明,HMGA2上調了CRC細胞中CCL2的表達,并以Stat3依賴的方式促進巨噬細胞的遷移。
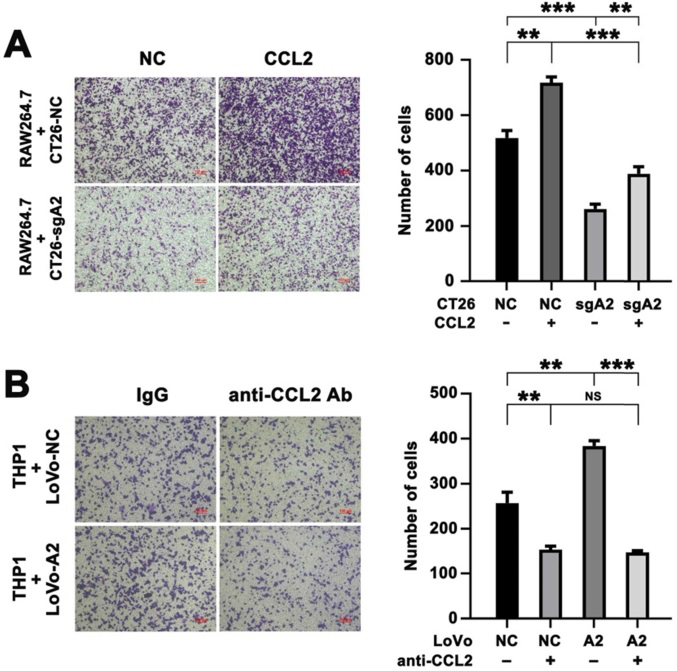
6)CRC細胞中HMGA2過表達導致巨噬細胞遷移的增加依賴于CCL2
為了研究CCL2在巨噬細胞招募中的作用,我們用重組小鼠CCL2處理CT26-NC和CT26-sgA2細胞,然后與RAW264.7細胞共培養。敲除Hmga2抑制了RAW264.7細胞的遷移潛能,而重組CCL2蛋白處理增強了其遷移潛能(圖6A)。此外,我們用中和性抗CCL2抗體預孵育LoVo細胞,然后使用Transwell共培養體系評估THP1細胞的遷移能力。如圖6B所示,我們的結果表明,HMGA2過表達促進THP1細胞的遷移,而抗CCL2抗體的處理則使THP1細胞遷移消失,說明HMGA2和CCL2在巨噬細胞招募中起著至關重要的作用。我們還發現,中和CCL2后,與LoVo-A2共培養的THP1遷移誘導被取消。這些數據表明,HMGA2過表達以CCL2依賴的方式促進巨噬細胞的遷移。
7)人結直腸癌標本中HMGA2和CD68表達的臨床意義
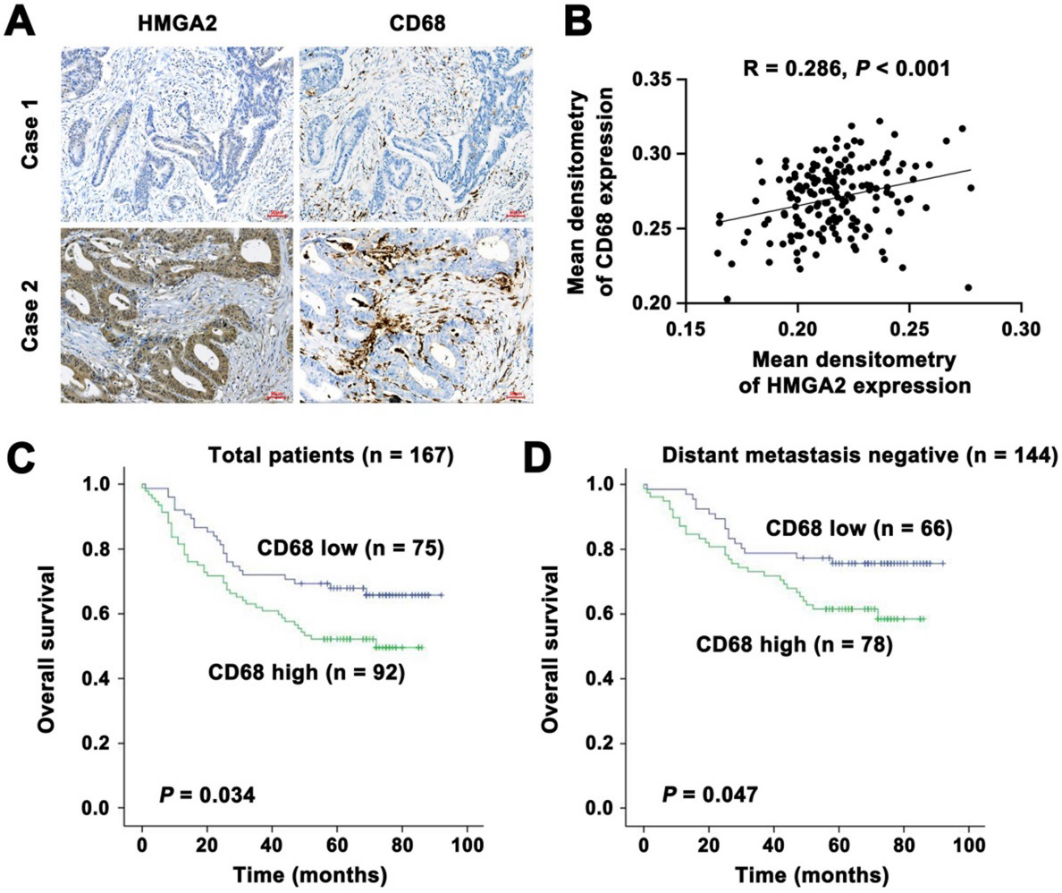
有報道稱CD68是巨噬細胞的免疫組化染色標記物。為了研究HMGA2水平與CRC患者巨噬細胞浸潤的關系,我們采用免疫組化染色對167例人類CRC標本中HMGA2和CD68的表達進行了評估。結果表明,腫瘤細胞中HMGA2的表達與間質中CD68的表達有很強的相關性。如圖7A-B所示,我們發現HMGA2與CD68表達呈正相關趨勢。接下來,我們進行Kaplan-Meier生存分析來評估CD68作為CRC預后標志物的潛在價值。如圖7C所示,在所有患者中,間質中CD68高表達與患者生存期降低相關。當分層到遠處轉移陽性和陰性亞組時,我們發現CD68高表達患者的總生存期較差,而CD68低表達患者的總生存期較好(圖7D)。這些結果提示間質CD68可作為臨床預后的標志和預測因子,提示其在結直腸癌中的臨床意義。
結論:CRC細胞中過表達HMGA2通過上調Sat3介導的CCL2分泌,促進巨噬細胞招募和M2極化,從而促進CRC腫瘤免疫抑制。我們的研究揭示了HMGA2在免疫抑制微環境形成中的一種新的促癌作用。
參考文獻:
Xin Wang, Jian Wang, Jiahui Zhao, Hao Wang, Jing Chen, Jingjing Wu. HMGA2 facilitates colorectal cancer progression via STAT3-mediated tumor-associated macrophage recruitment. Theranostics 2022; 12(2): 963-975. doi:10.7150/thno.65411.