






在結直腸癌中, CircHERC4作為一種新的致癌驅動因子,通過miR - 5565p /CTBP2/E- cadherin軸促進腫瘤轉移
結直腸癌(Colorectal cancer,CRC)患者死亡的主要原因是轉移。越來越多的證據表明,CircRNA在癌癥的起始和發展中發揮著關鍵作用。然而,CircRNAs協同癌癥轉移的潛在分子機制仍不明確。本研究探討CircHERC4在CRC中的作用機制。本文于2021年10月發表在《Journal of Hematology & Oncology》,IF= 11.059。
本文技術路線:

主要結果
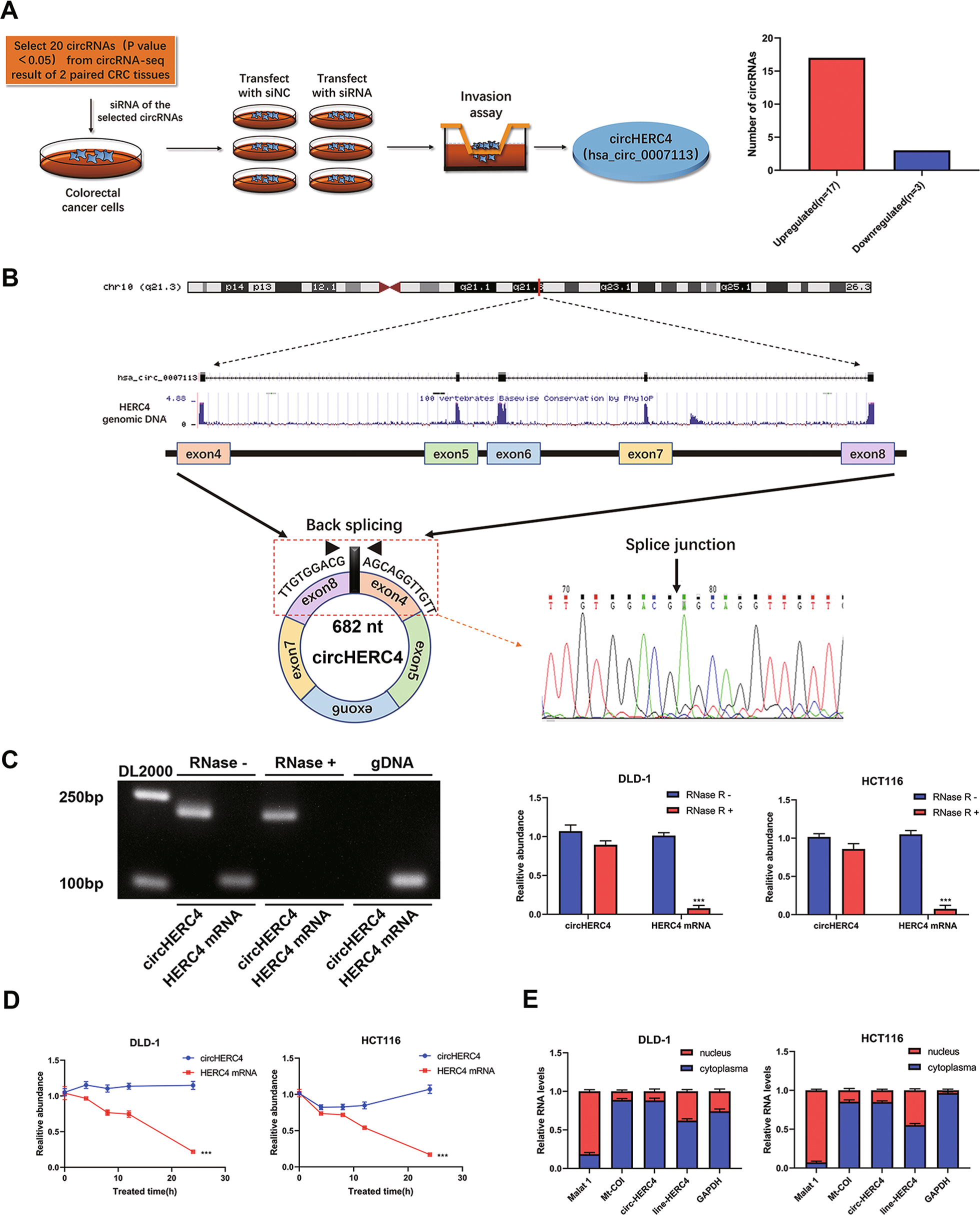
1. CircHERC4在CRC中的識別與特征
首先,選擇兩對CRC組織和癌旁組織進行RNAseq,分析CircRNAs的表達譜,發現CRC組織中20個差異表達的CircRNA (FC≥10,P < 0.05)。為了篩選與轉移相關的CircRNA,作者將這些被篩選的CircRNA中進行siRNA轉染到CRC細胞中,并采用Transwell侵襲試驗篩選出具有功能的環狀RNA。結果發現CircHERC4 (has_Circ_0007113) 是CRC中過表達的最顯著的驅動轉移的CircRNA (Fig 1A)。CircBase報道CircHERC4來自HERC4基因,由4-8外顯子(682 bp)頭尾剪接組成。于是作者設計了一個分歧引物來擴增HERC4剪接形式,并通過Sanger測序證實CircHERC4中存在剪接連接(Fig 1B)。在DLD-1細胞中,Circ HERC4只能在cDNA中擴增,而不能在gDNA中擴增,這排除了由基因組重排引起的可能(Fig 1C)。此外,通過RNase R處理和放線菌素D實驗檢測CircHERC4的穩定性。RNase R和放線菌素D處理后,HERC4 mRNA的水平顯著下調,而CircHERC4 mRNA水平保持不變(Fig 1C,D)。此外,通過核質分離后的qRT-PCR,發現CircHERC4主要位于細胞質中(Fig 1E)。
2. CircHERC4 mRNA的過表達與CRC患者預后不良相關
為了進一步明確CircHERC4在CRC中的致癌表型,采用qRT-PCR檢測了120對組織樣本中CircHERC4的表達情況。結果表明,CircHER4表達水平在CRC患者中明顯升高(Fig 2A)。此外,CircHERC4高表達與淋巴轉移(Fig 2B)和遠處轉移(主要是肝轉移)呈正相關(Fig 2C)。進一步探索CircHERC4的過表達是否由宿主HERC4 mRNA的上調引起CRC。然而,在120個CRC組織中沒有發現HERC4 mRNA顯著增加(Fig 2D)。同樣的,HERC4 mRNA與淋巴轉移和肝轉移也無相關性((Fig 2E,F)。因此,作者推測CircHERC4的過表達在CRC中是轉錄后被調控的。此外,Kaplan-Meier生存分析顯示,CircHERC4高表達的患者總生存期較差(Fig 2G)。HERC4 mRNA表達水平與患者預后無相關性(Fig 2H)。綜上所述,這些結果表明,CircHERC4(而非其宿主HERC4 mRNA)在CRC組織中顯著上調,而更高的CircHERC4表達可能意味著預后不良。
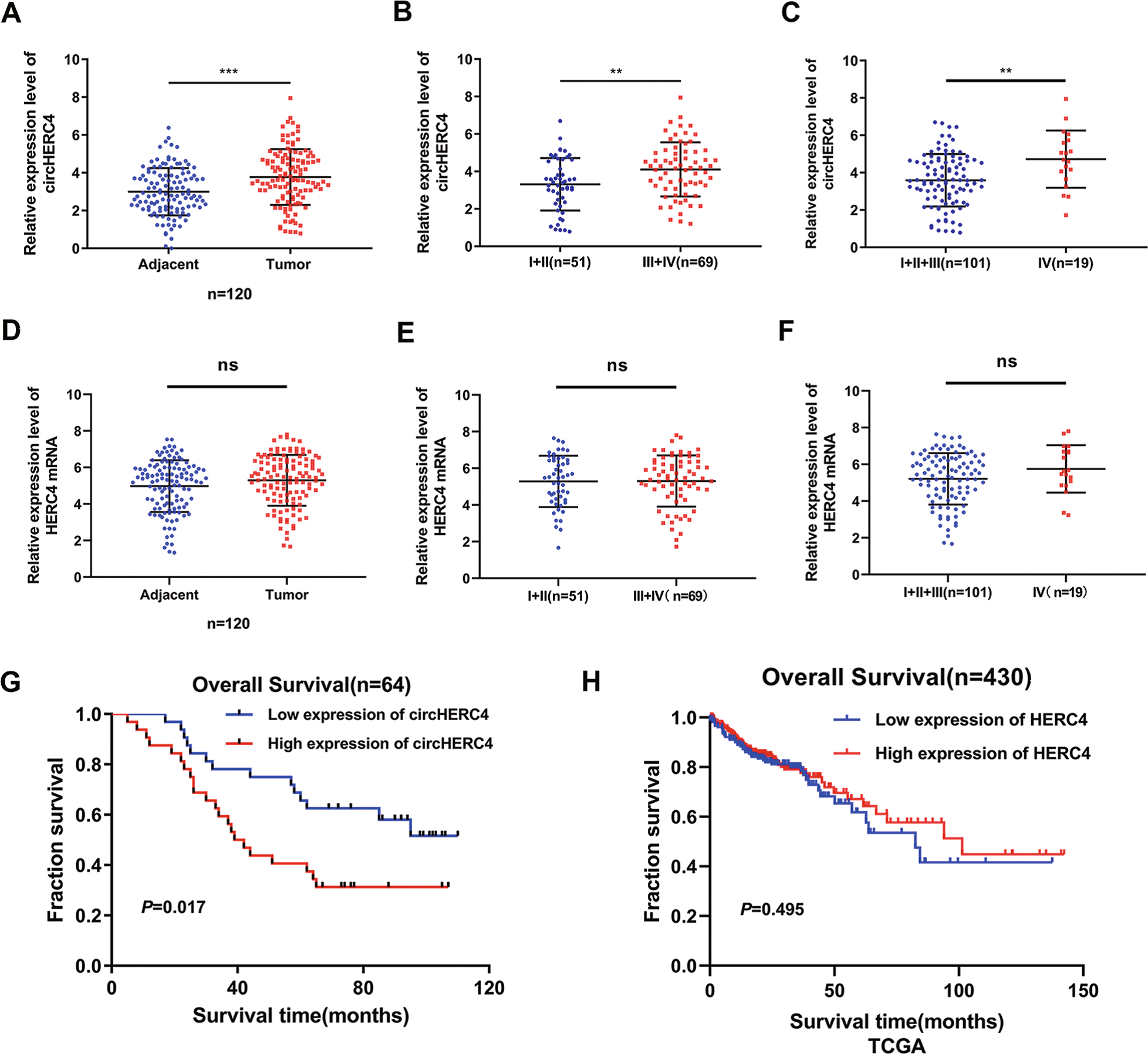
Fig 2 CircHERC4在CRC組織中過表達并與轉移相關
3. CircHERC4作為一種致癌基因,在體外和體內促進CRC的增殖、遷移和侵襲
為了確定CircHERC4的功能作用,作者檢測了CircHERC4在人正常結腸上皮細胞(HCoEpic)和CRC細胞 (HCT116,DLD-1,HT29,LoVo和SW480) 中的表達。作者發現與正常結腸上皮細胞HCoEpic細胞相比,CircHERC4在CRC細胞中表達明顯上調,CircHERC4在DLD-1和HCT116細胞中表達相對較高,而在SW480細胞中表達相對較低(Fig 3A)。因此,作者選擇HCT116、DLD-1和SW480細胞進行進一步研究。接下來針對CircHERC4后剪接序列設計了siRNAs(Fig 3B)。用CircHERC4 siRNA轉染DLD-1和HCT116細胞,成功地沉默了CircHERC4 (Fig 3C)。CCK-8實驗顯示,沉默CircHERC4可顯著抑制DLD-1和HCT116細胞的活力(Fig 3D)。此外,集落形成實驗顯示,與陰性對照相比,CircHERC4下調組的DLD-1和HCT116細胞的增殖能力也受到抑制(Fig 3E)。此外,transwell實驗顯示,沉默CircHERC4可以抑制DLD-1和HCT116細胞的遷移和侵襲能力(Fig 3F)。而成功構建過表達CircHERC4質粒并轉染到SW480細胞中,發現CircHERC4明顯增加(Fig 3G),提示CircHERC4上調可顯著促進體外CRC細胞的增殖、遷移和侵襲能力(Fig 3H–J)。
此外,異種移植和轉移小鼠模型也支持CircHERC4在體內發揮致癌作用的觀點。在皮下腫瘤模型中,CircHERC4沉默(sh-CircHERC4-1和shCircHERC4-2)和陰性對照(sh-NC) HCT116細胞通過皮下注射到BALB/c裸鼠中。每7天測量腫瘤體積,并讓腫瘤生長35天。結果顯示,與陰性對照組相比,sh-CircHERC4組的腫瘤生長速度和腫瘤重量明顯受到抑制(Fig 3K–M)。將上述三組HCT116細胞轉移到腫瘤小鼠模型中,小鼠生長60天后,處死小鼠,取肺、肝進行HE染色,檢測轉移灶的形成。發現肺和肝臟均有轉移性結節(Fig 3N)。與陰性對照組相比,sh-CircHERC4組肺、肝轉移的發生率顯著降低(Fig 3N)。HE染色進一步顯示sh-NC組轉移結節面積極高,而sh-CircHERC4組轉移結節面積顯著降低(Fig 3O)。以上數據證實了CircHERC4在體外和體內均能促進CRC的進展,特別是轉移。
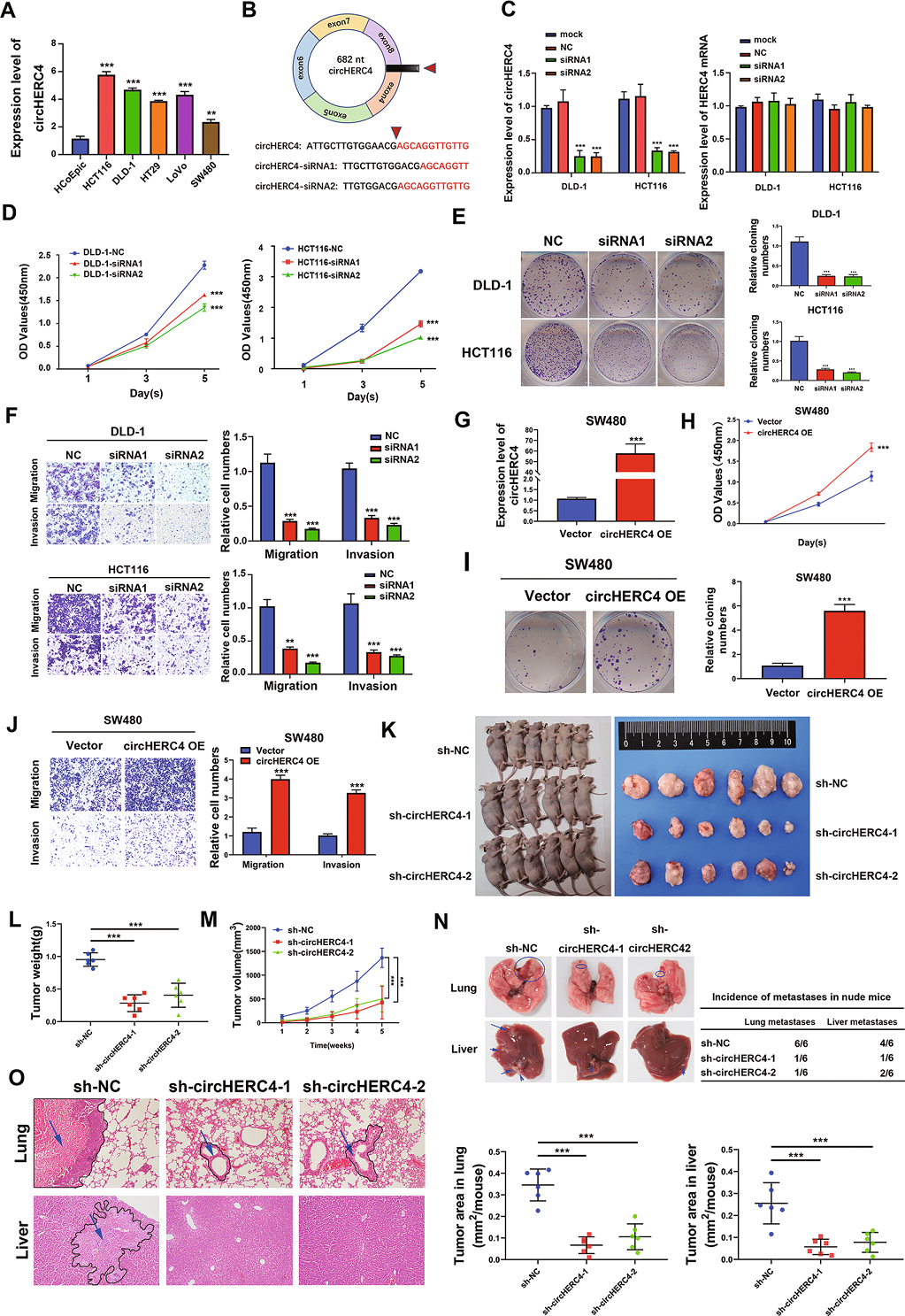
Fig3 CircHERC4作為一種致癌基因,在體外和體內調控增殖、遷移和侵襲
4. CircHERC4直接與CRC細胞中的miR - 5565p結合
CircHERC4主要位于CRC細胞的細胞質中,提示其可能具有轉錄后功能。因此,作者通過StarBase和Circinteractome預測其潛在的結合miRNA。作者初步篩選出了36miRNAs。為了確定這些miRNA是否能與CircHERC4結合,通過miRNA模擬物進行了熒光素酶篩選。作者構建了一個CircHERC4片段,并將其插入psiCHECK-2質粒中熒光素酶報告基因的下游。然后將miRNA模擬物與luc-CircHERC4報告質粒共轉染293T細胞。與對照相比,其中7個miRNAs可以使熒光素酶活性降低至少40%,在雙熒光素酶報告實驗中,miR-556-5p對熒光素酶活性的抑制作用最強 (Fig 4A)。為了證實這一預測,作者設計了靶向連接位點的生物素化CircHERC4探針和scrambled oligo probes探針并用于在DLD-1細胞中進行RNA下拉試驗。RNA下拉實驗后的qPCR結果顯示,CircHERC4 mRNA顯著富集,而宿主HERC4 mRNA則不顯著富集。通過qPCR檢測CircHERC4拉下部分中所有具有熒光素酶活性抑制作用的7個miRNAs。miR-556-5p在CircHERC4下拉部分的富集顯著高于NC組和其他miRNAs (Fig 4B)。這些結果提示miR-556-5p能直接與CircHERC4結合,并參與CircHERC4在CRC中的功能。

Fig 4 CircHERC4通過與miR-556-5p相互作用促進CRC細胞增殖、遷移和侵襲
5. miR -5556- 5p的抑制介導了CircHERC4的致癌功能,并激活了CRC中的Notch信號通路
由于目前尚無關于miR-556-5p在CRC的研究,作者首先通過qPCR檢測證實了miR-556-5p在臨床CRC樣本中的表達模式。結果顯示,與癌旁組織相比,miR-556-5p在CRC組織中下調(Fig 4C)。作者進一步分析了miR-556-5p在有無轉移的CRC患者中的表達情況,結果顯示miR-556-5p在有淋巴轉移(Fig 4D)或遠處轉移患者中表達水平明顯降低(Fig 4D,E)。為了探究miR-556-5p的功能作用,作者將miR-556-5p模擬物轉染到DLD-1和HCT116細胞中。所有結果顯示,miR-556-5p可顯著抑制CRC細胞的增殖、遷移和侵襲能力(Fig 4F-H)。此外,通過挽救實驗在SW480中共轉染miR- 556-5p模擬物和CircHERC4過表達載體。結果發現,miR-556-5p模擬物可以部分恢復CircHERC4過表達增強SW480細胞的增殖、遷移和侵襲能力(Fig 4I-K)。綜上所述, miR-556-5p具有抗癌作用,并通過直接結合在CRC中部分介導CircHERC4的致癌功能。
然而,可能受到CircHERC4/miR-556-5p軸的影響的下游信號通路尚不清楚。因此,作者在沉默CircHERC4或過表達miR-556-5p后進行了RNA測序。根據維恩圖顯示,作者篩選出了9109個在CircHERC4沉默和miR-556-5p過表達的DLD-1細胞中顯著改變的轉錄本,KEGG通路富集分析表明,這9109個轉錄本富集在許多腫瘤相關信號通路中,尤其是被廣泛認為是大腸癌發生激活因子的Notch信號通路(Fig 5B)。作者假設CircHERC4可能通過激活Notch信號通路促進細胞轉移,而Notch信號通路可能被miR-556-5p阻斷。
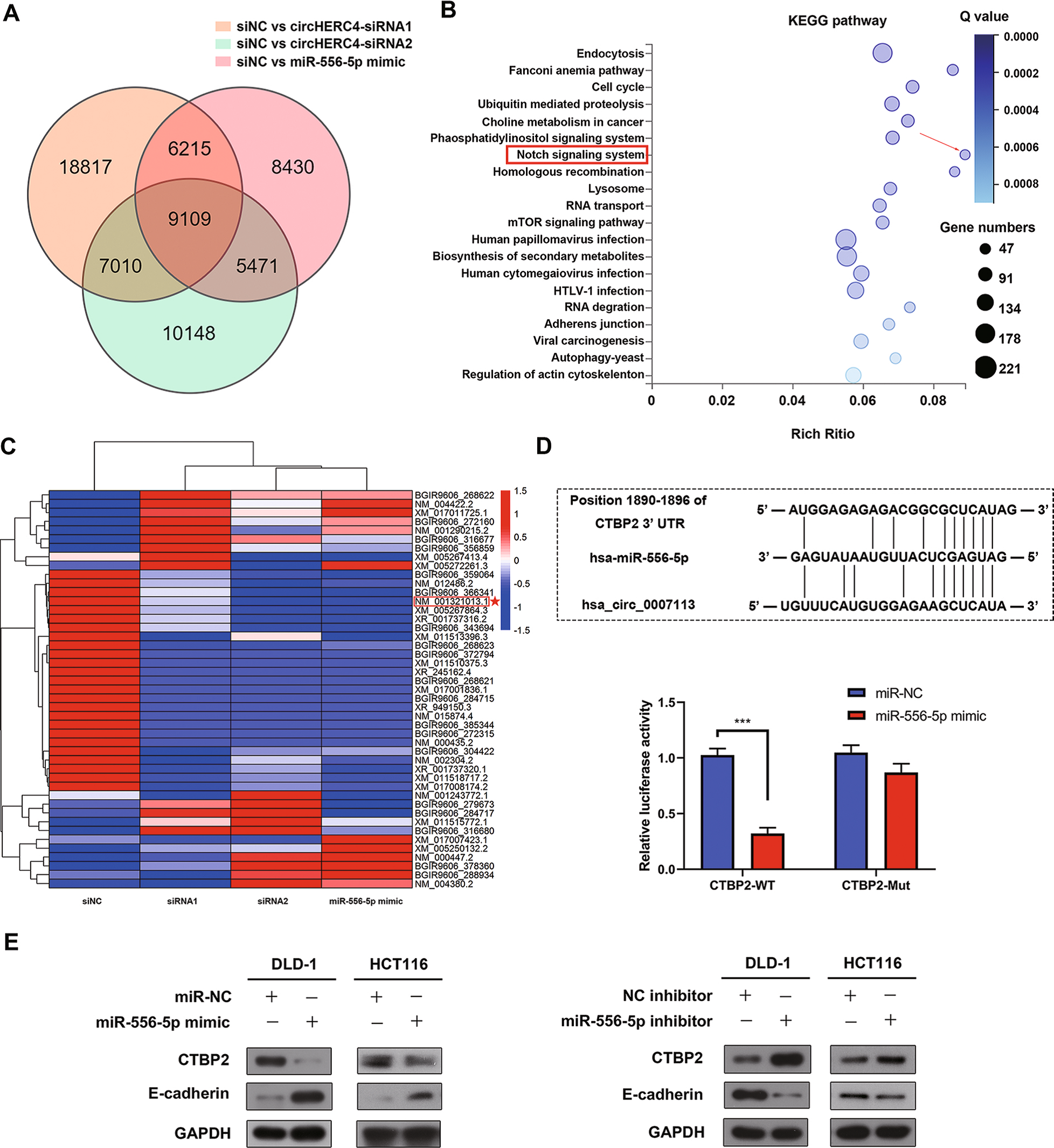
Fig 5 CTBP2是miR-556-5p的靶基因
6. CTBP2是miR - 5565p的靶點,通過抑制結直腸癌中E - cadherin的表達發揮致癌作用
作者進一步探討了miR-556-5p靶向的Notch信號通路的效應因子。熱圖顯示,Notch信號通路中的大部分基因通過沉默CircHERC4和過表達miR-556-5p而下調(Fig 5C)。結合生物信息學分析,CTBP2被預測為miR-556-5p的靶點(Fig 5D)。為了進一步證實這一假設,作者進行了熒光素酶實驗來驗證miR-556-5p和CTBP2之間的相互作用。將含有miR-556-5p野生型和突變型結合位點的CTBP2 3’UTR片段克隆到熒光素酶報告質粒psiCHECK2中,并與miR-556-5p 模擬物或miR-NC模擬物共轉染到DLD-1細胞中。結果表明過表達miR-556-5p顯著降低了包含野生型結合位點載體的熒光素酶活性,而非包含突變型結合位點載體的熒光素酶活性(Fig 5D)。此外,作者通過WB實驗證明,miR-556-5p可以顯著抑制CTBP2的蛋白水平,但卻挽救了CTBP2[26]的靶基因E-cadherin的表達(Fig 5E)。
CTBP2被認為是結直腸癌中腫瘤發展的一個重要方面。作者分析了GEPIA數據庫,發現CTBP2蛋白在結直腸癌組織中中位表達水平高于癌旁正常組織(Fig 6A)。 接下來,研究了120例CRC樣本中CTBP2 mRNA的表達,在免疫組化中觀察到了CTBP2在CRC中高表達(Fig 6B)。此外,生存分析評估也發現了CTBP2高表達與CRC患者較差的生存概率顯著相關(Fig 6C)。作者構建并轉染CTBP2 siRNA到DLD-1和HCT116細胞中,通過WB實驗證實E-cadherin蛋白表達水平下降(Fig 6D)。與對照組相比,沉默CTBP2顯著抑制DLD-1和HCT116細胞增殖、遷移和侵襲能力的表達(Fig 6E-G)。這些結果證明了CTBP2是miR-556-5p的靶基因,其過表達通過抑制E-cadherin介導CRC腫瘤轉移。

Fig 6 CTBP2在CRC組織中過表達,可促進CRC細胞的發展
7. 沉默CTBP2可以消除過表達CircHERC4引起的作用
作者已經證實CircHERC4可以阻斷miR-556-5p,于是進一步驗證CircHERC4是否能調控miR-556-5p靶蛋白CTBP2的表達。轉染lv - CircHERC4– shRNA到DLD-1和HCT116細胞中。結果發現,CircHERC4基因敲除后,CTBP2蛋白水平顯著降低。相反,CTBP2的下游靶點E-cadherin顯著增加(Fig 6H)。免疫組化染色檢測sh-NC、sh-CircHERC4-1和sh-CircHERC4-2組HCT116細胞中CTBP2、E-cadherin和Ki67的表達。發現CircHERC4沉默后E-cadherin的表達水平被誘導,Ki-67在急劇下調(Fig 6i)。為了進一步研究CircHERC4是否通過誘導CTBP2的表達來產生致瘤性,作者進行了一項挽救實驗來研究CircHERC4和CTBP2之間的功能相互作用。結果發現,與感染空載或siNC模擬物的SW480相比,轉染CTBP2 siRNA模擬物的過表達CircHERC4的細胞增殖、遷移和侵襲能力明顯下降(Fig 6J-I),提示CircHERC4的致癌功能可以通過消除CTBP2的表達而部分逆轉。WB結果顯示,CTBP2可以部分挽救CircHERC4對E-cadherin表達的影響(Fig 6m)。
此外,根據CRC樣本的臨床分期,對10個高級別和10個低級別CRC樣本進行FISH檢查,以證實CircHERC4/miR-556-5p/CTBP2軸之間的關系。結果發現CircHERC4和CTBP2在高級別CRC組織中過表達,而miR-556-5p在低級別組織中含量較高(Fig 7A)。最后,進行了包含四組體內挽救實驗(vector,CircHERC4 OE, CircHERC4 OE + CTBP2 siRNA, CircHERC4 OE + miR-556-5p mimic)。異種皮下移植結果表明,CircHERC4過表達可加速腫瘤生長,注射CTBP2 siRNA或miR-556-5p mimic后,這種作用可被抑制(Fig 7B-D)。
此外,免疫組化實驗表明, 過表達CircHERC4可上調CTBP2和Ki67,下調E-ca的表達;這種效果可以通過抑制CTBP2或miR-556-5p過表達挽救(Fig 7E)。在尾靜脈注射小鼠模型中,上調的CircHERC4促進了肺和肝轉移結節的形成,當CTBP2下調或miR-556-5p上調時,CircHERC4誘導的致癌作用被阻斷(Fig 7F-G)。總的來說,這些結果證明CTBP2可以促進CRC的進展,并且在體內和體外都受到CircHERC4/miR-556-5p軸的調控。
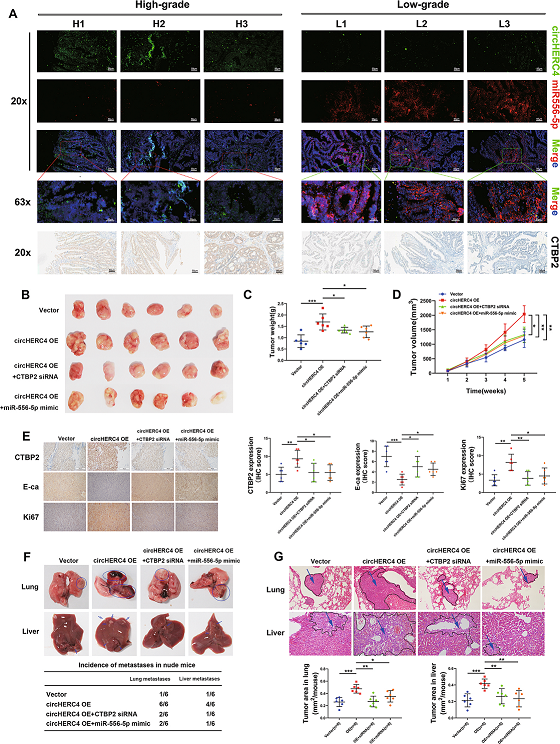
Fig7 CTBP2受circHERC4/miR-556-5p相互作用的調控
綜上所述,本研究發現了一個之前未被發現的致癌驅動因子CircHERC4。CircHERC4在CRC組織中升高,并與CRC的增殖、遷移和侵襲相關。此外,CircHERC4在CRC患者中的高表達與轉移和較差的生存率呈正相關。作者提出了CircHERC4可以阻斷miR-556-5p對CTBP2的抑制活性的機制;因此,沉默CircHERC4可以下調表達CTBP2同時活化E-cadherin(Fig 8)。

Fig 8闡明circHERC4通過miR-556-5p/CTBP2/E-cadherin信號通路促進CRC發病和轉移的機制
參考文獻:
He, J., et al., Circular RNA circHERC4 as a novel oncogenic driver to promote tumor metastasis via the miR-556-5p/CTBP2/E-cadherin axis in colorectal cancer. J Hematol Oncol, 2021. 14(1): p. 194.