






順式作用lnc-Cxcl2抑制中性粒細胞介導的病毒感染性肺炎
病毒感染過程中上皮細胞產生的趨化因子對于中性粒細胞的招募非常重要,適當的調節(jié)趨化因子產生對于限制炎癥和減緩隨后的組織損傷非常重要。在病毒感染和炎癥過程中,上皮細胞lncRNA、RNA結合蛋白的表達及其功能相互作用尚不清楚。本文發(fā)現lnc-Cxcl2順式作用通過抑制病毒感染中上皮細胞CXCL2的表達來抑制中性粒細胞介導的肺炎。該于2021年10月發(fā)表在《Proceedings of the National Academy of Sciences of the United States》IF:9.58期刊上。
技術路線:

主要實驗結果:
1、lnc-Cxcl2的表達在小鼠肺上皮細胞核中上調表達以響應病毒感染
首先,作者通過文獻確定了趨化因子CXCL2為研究目前,然后通過RNA測序篩選到順式調節(jié)CXCL2的lncRNA,將其命名為lnc-Cxcl2。首先檢測了lnc-Cxcl2的表達,如圖1A-D所示,和測序結果一致,lnc-Cxcl2在肺上皮細胞系中的表達響應病毒感染,并和CXCL2的mRNA表達趨勢一致。并且用WB檢測到了其轉錄本的表達(圖1E)。原位雜交顯示隨著病毒感染延長,lnc-Cxcl2在核中的組分逐漸增加(圖1F)。NF-κB inhibitor處理后,CXCL2的mRNA表達和lnc-Cxcl2的表達都顯著下降了(圖1H-G)。表明lnc-Cxcl2是一個核lncRNA,通過NF-κB信號通路響應病毒感染。
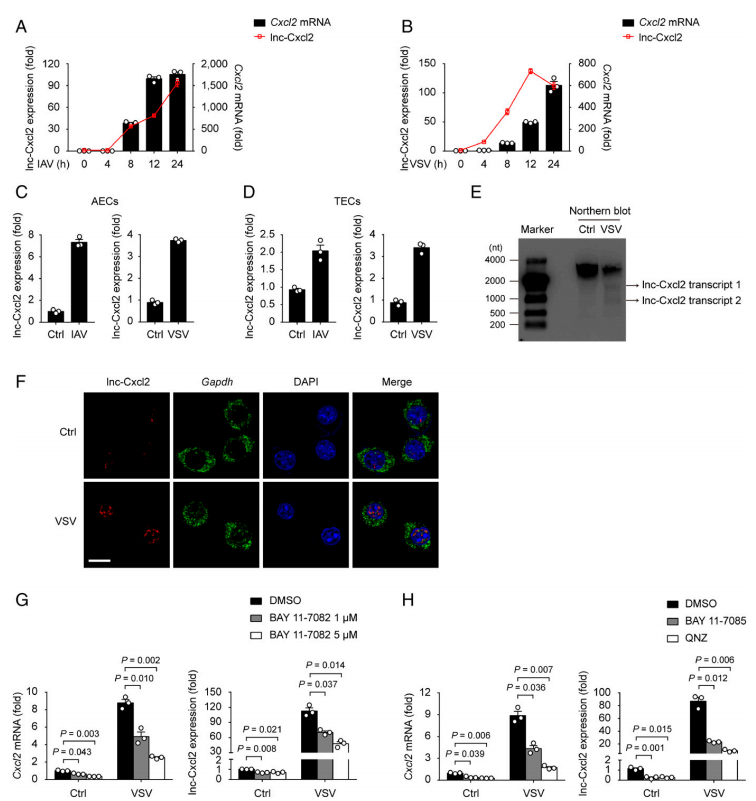
圖1 lnc-Cxcl2在小鼠肺上皮細胞核中表達增加,以應對病毒感染
2、lnc-Cxcl2選擇性抑制病毒感染期肺上皮細胞CXCL12的表達
對lnc-Cxcl2敲除和對照的細胞進行病毒感染,然后轉錄組測序,發(fā)現21個基因在敲除組異常表達(圖2A),其中CXCL2上調表達(圖2B)。和轉錄組測序一致,我們發(fā)現Cxcl2的表達和分泌型CXCL2的表達在lnc-Cxcl2敲除組顯著升高(圖2C-D)。并且lnc-Cxcl2缺陷增加了IAV和VSV感染的AECs和TECs細胞中的Cxcl2表達,但在AMs中沒有(圖2E)。因此, lnc-Cxcl2在病毒感染時選擇性地抑制小鼠肺上皮細胞中Cxcl2的表達。

圖2 lnc-Cxcl2選擇性抑制病毒感染時肺上皮細胞中Cxcl2的表達
lnc-Cxcl2?/?小鼠在外觀、體重和行為上與lnc-Cxcl2+/+同窩小鼠幾乎沒有區(qū)別。并且lnc-Cxcl2+/+和lnc-Cxcl2?/?小鼠脾臟免疫細胞包括T細胞、B細胞、自然殺傷細胞、中性粒細胞、巨噬細胞和樹突狀細胞的比例也相似,提示lnc-Cxcl2缺乏并不影響小鼠免疫細胞的發(fā)育。所以,用IAV鼻腔感染這些小鼠,以肺上皮細胞為靶點感染并誘導肺炎癥和組織損傷。結果發(fā)現,與lnc-Cxcl2+/+小鼠相比,lnc-Cxcl2?/?小鼠在IAV感染后,體重下降更多,肺中Cxcl2的表達增加,但不包括其他促炎細胞因子,如Ccl2、Ifnb1或Tnf (圖3A-B)。此外,lnc-Cxcl2?/?小鼠支氣管肺泡灌洗液(BALF)中CXCL2水平和中性粒細胞數均高于lnc-Cxcl2+/+小鼠(圖3C-D)。與此一致的是,IAV感染后lnc-Cxcl2?/?小鼠與lnc-Cxcl2+/+小鼠相比,肺中性粒細胞浸潤更嚴重,炎癥更嚴重,組織損傷更嚴重(圖3E-F)。表明lnc-Cxcl2?/?小鼠中由于CXCL2的表達增高所以更多的中性粒細胞被募集至肺部,加重病情。表明lnc-Cxcl2通過抑制CXCL2表達和中性粒細胞募集抑制流感病毒誘導的體內肺炎癥。
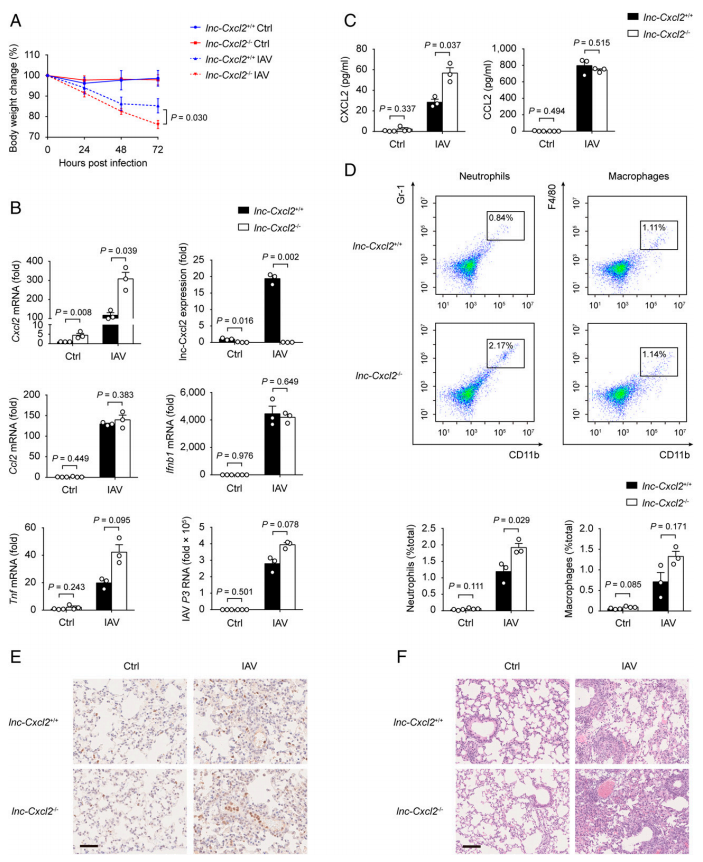
圖3 lnc-Cxcl2在體內抑制流感病毒誘導的肺炎癥
4、lnc-Cxcl2以順式作用結合并維持Cxcl2啟動子的染色質抑制狀態(tài)
為了探究lnc-Cxcl2如何抑制Cxcl2的,首先作者檢測了lnc-Cxcl2對轉錄因子的調控,發(fā)現核定位轉錄因子P65, P50, STAT1, STAT3或c-JUN的表達都不受lnc-Cxcl2的調控,所以猜測lnc-Cxcl2是以順式作用的方式調控Cxcl2。作者發(fā)現lnc-Cxcl2缺陷增加了Cxcl2 pre-mRNA水平以應對病毒感染(圖4A),Cxcl2啟動子的染色質可訪問性(以DNase敏感性表示)也增加了,以響應病毒感染(圖4 B)。一致地,Cxcl2啟動子中H3K4me3和CREB結合蛋白(CBP)的水平,是兩種活性轉錄標記,在lnc-Cxcl2敲除細胞中增加(圖4C-D)。Cxcl2啟動子的總RNA聚合酶和磷酸化RNA聚合酶II水平也在lnc-Cxcl2敲除細胞中增加(圖4E-F),提示lnc-Cxcl2抑制Cxcl2啟動子的染色質可及性和Cxcl2的轉錄。
為了探究lnc-Cxcl2如何調控Cxcl2啟動子染色質可及性,使用antisense和lnc-Cxcl2探針分離lnc-Cxcl2結合的染色質,發(fā)現探針可以成功捕獲lnc-Cxcl2,特別是在VSV干擾后,并且Cxcl2的啟動子兩個κB區(qū)域顯著富集lnc-Cxcl2探針。表明在病毒感染期間,lnc-Cxcl2順式結合Cxcl2啟動子并維持Cxcl2啟動子染色質的抑制狀態(tài)。
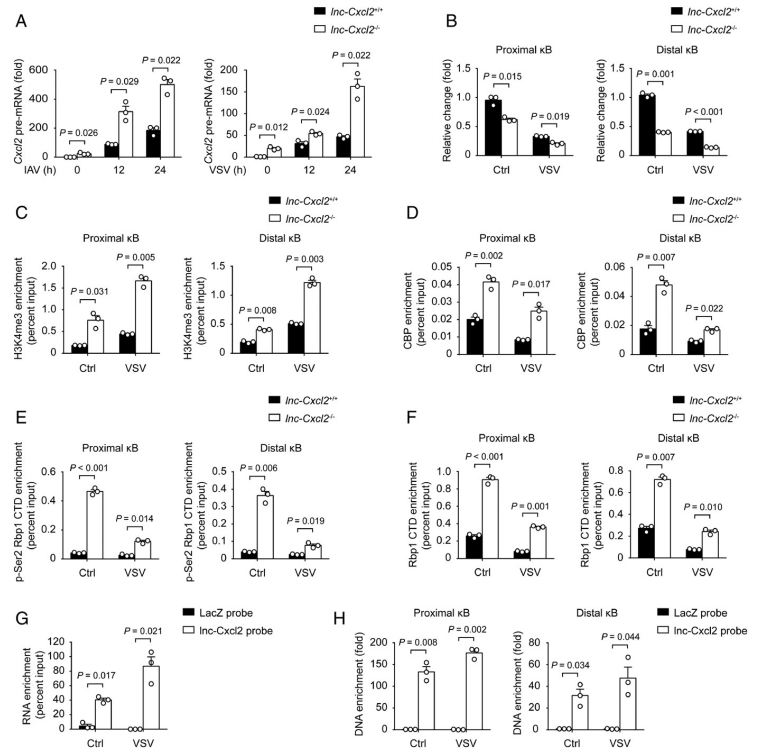
圖4 lnc-Cxcl2以順式結合并維持Cxcl2啟動子受抑制的染色質狀態(tài)
5、lnc-Cxcl2通過核糖核蛋白La抑制Cxcl2表達
接下來探究lnc-Cxcl2是否通過其相互作用的蛋白抑制染色質的可及性。使用RNA pull-down和MS鑒定與lnc-Cxcl2相互作用的核蛋白,得到核糖核蛋白La。為了驗證lnc-Cxcl2通過與La相互作用抑制Cxcl2轉錄的假設,首先證實lnc-Cxcl2與La之間的相互作用(圖5A-B)。構建La的截短突變體,發(fā)現只有包含第二個RRM的La片段與lnc-Cxcl2結合,表明La通過RRM2與lnc-Cxcl2相互作用(圖5C)。為了研究La在調節(jié)Cxcl2表達中的作用,構建了La缺陷(Ssb?/?)的MLE-12細胞。發(fā)現La缺失不僅增加了Cxcl2的表達,而且還增加了其他趨化因子和促炎細胞因子的表達,如Ccl2、Ifnb1和Tnf,以應對病毒感染(圖5D)。然后,研究了La如何抑制這些細胞因子的表達,發(fā)現La缺失增加了Cxcl2和Ccl2啟動子的染色質可及性(圖5E)。總之,雖然La抑制許多細胞因子的表達,但lnc-Cxcl2可以結合Cxcl2啟動子,在病毒感染過程中通過核糖核蛋白La抑制Cxcl2順式表達。

圖5 lnc-Cxcl2通過核糖核蛋白La抑制Cxcl2表達
6、lnc-CXCL2-4-1抑制人肺上皮細胞免疫細胞因子表達
雖然lncRNA在不同物種之間被認為是低保守的,但在人類細胞中是否也存在類似的機制。通過搜索lncRNA數據庫,在人類CXCL2的下游發(fā)現了4個與CXCL2相關的轉錄本,分別為lnc-CXCL2-1-1、lnc-CXCL2-2-1、lnc-CXCL2-3-1、lnc-CXCL2-4-1。其中,lnc-CXCL2-3-1預測有編碼框,因此只有其他3個轉錄本被認為是ncRNA,然而,這三種lncRNA的具體功能尚未被闡明。因此,首先檢測了這些lncRNA在A549人肺上皮細胞中的表達,發(fā)現IAV感染后lnc-CXCL2-1-1的表達顯著增加,而lnc-CXCL2-1-1和lnc-CXCL2-2-1的表達僅輕微增加(圖6A)。然后檢測lnc-CXCL2- 4-1是否參與了趨化因子表達的調控。通過轉錄組分析發(fā)現,IAV感染后lnc-CXCL2-4-1沉默的A549細胞中有364個基因表達上調,32個基因表達下調(圖6B)。對這些差異表達基因的GO分析表明,它們的分子功能豐富于細胞因子活性,包括趨化因子、干擾素和其他促炎細胞因子(圖6C-D)。進一步證實沉默lnc-CXCL2-4-1確實增加了IAV感染后A549細胞中CXCL2和CXCL8的表達。此外,lnc-CXCL2-4-1與La相互作用,沉默La的表達增加了IAV感染后A549細胞中CXCL2和CXCL8的表達(圖6 F和G),表明lnc-CXCL2-4-1抑制細胞因子的表達。綜上所述,這些結果表明,不同物種的宿主通過增加一種lncRNA的表達來抑制病毒誘導的肺炎癥,該lncRNA可以反饋抑制肺上皮細胞中趨化因子的產生。

圖6 lnc-CXCL2-4-1抑制人肺上皮細胞中細胞因子的表達
參考文獻:
Liu Shuo., Liu Jiaqi., Yang Xue., Jiang Minghong., Wang Qingqing., Zhang Lianfeng., Ma Yuanwu., Shen Zhongyang., Tian Zhigang., Cao Xuetao.(2021). Cis-acting lnc-Cxcl2 restrains neutrophil-mediated lung inflammation by inhibiting epithelial cell CXCL2 expression in virus infection. Proc Natl Acad Sci U S A, 118(41), undefined. doi:10.1073/pnas.2108276118