






M6A RNA甲基化介導(dǎo)的RMRP穩(wěn)定性導(dǎo)致非小細(xì)胞肺癌的增殖和進(jìn)展
在全球所有惡性腫瘤中,非小細(xì)胞肺癌(NSCLC)的死亡率最高。 lncRNA在腫瘤進(jìn)展中的作用是當(dāng)前的研究熱點(diǎn)。基于TCGA數(shù)據(jù)庫的綜合分析,我們發(fā)現(xiàn)RMRP是與NSCLC低生存率相關(guān)的最高上調(diào)的lncRNA之一。此外,m6A在RMRP內(nèi)高度富集,增強(qiáng)了其RNA穩(wěn)定性。體外和體內(nèi)實(shí)驗(yàn)表明,RMRP可促進(jìn)NSCLC細(xì)胞增殖、侵襲和遷移。在機(jī)制上,RMRP將YBX1招募到TGFBR1啟動(dòng)子區(qū)域,導(dǎo)致TGFBR1的轉(zhuǎn)錄上調(diào)。TGFBR1/SMAD2/SMAD3通路也受RMRP調(diào)控。此外,RMRP促進(jìn)了腫瘤干細(xì)胞特性和上皮間充質(zhì)轉(zhuǎn)化,從而促進(jìn)了對放療和順鉑的耐藥。臨床資料進(jìn)一步證實(shí)RMRP與TGFBR1呈正相關(guān)。我們的工作揭示了m6A RNA甲基化介導(dǎo)的RMRP穩(wěn)定性通過調(diào)控TGFBR1/SMAD2/SMAD3通路而導(dǎo)致NSCLC的增殖和進(jìn)展。本文于2021年10月發(fā)表在“Cell Death & Differentiation”( IF:15.828)期刊上。
技術(shù)路線

結(jié)果
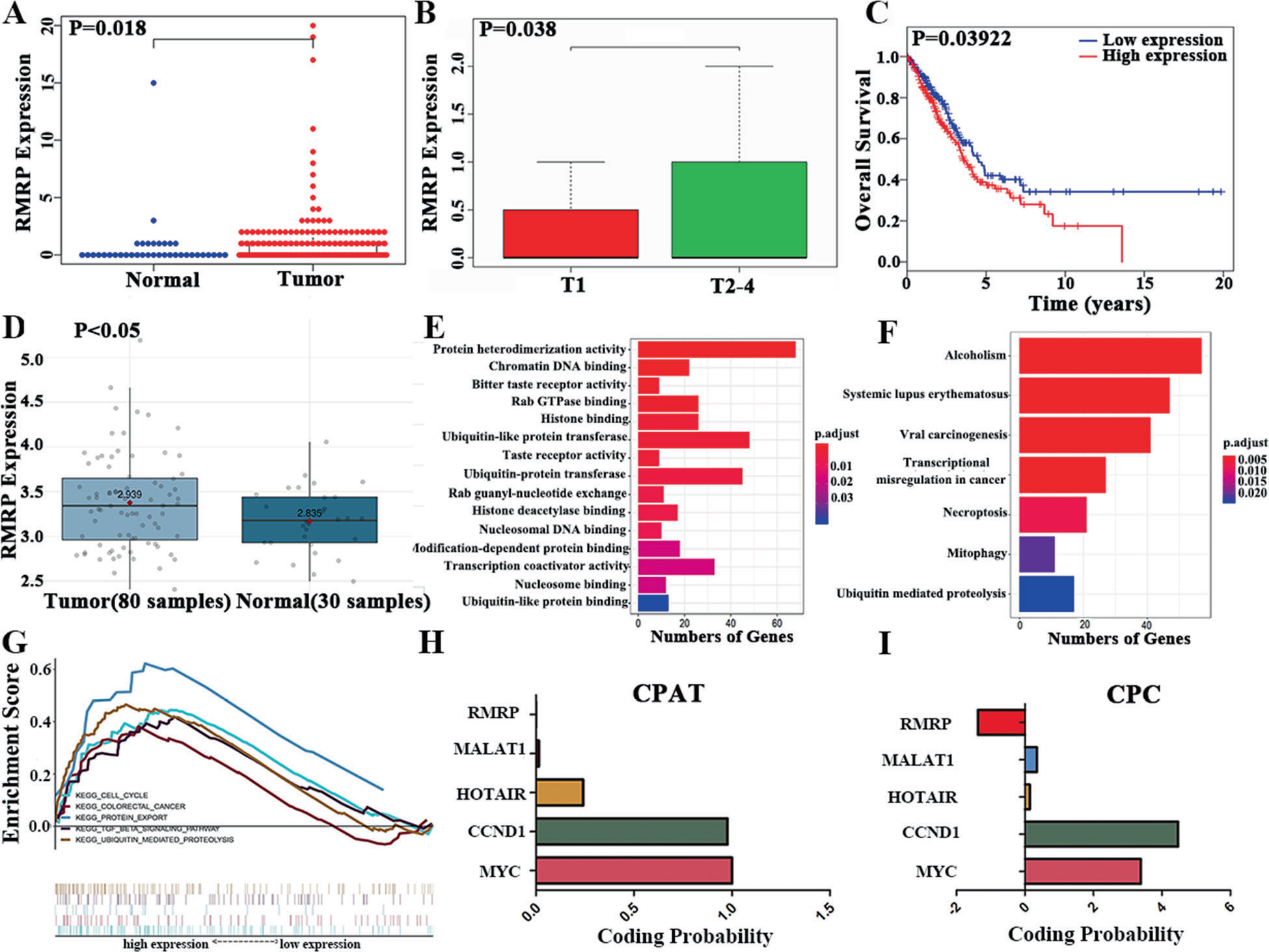
1)NSCLC的綜合分析顯示RMRP可能是一種生物標(biāo)志物
我們對TCGA肺腺癌(LUAD)數(shù)據(jù)庫進(jìn)行了綜合分析。與正常組織相比,RMRP在非小細(xì)胞肺癌組織中上調(diào)(圖1A)。高RMRP表達(dá)與晚期T期和較差的生存率相關(guān)(圖1B和C)。此外,在GSE43458中,與正常肺組織相比,非小細(xì)胞肺癌組織中RMRP表達(dá)上調(diào),如圖GEO數(shù)據(jù)庫(圖1D)所示。GO結(jié)果顯示,RMRP在蛋白質(zhì)異二聚活性、染色質(zhì)DNA結(jié)合、苦味受體活性等方面發(fā)揮作用(圖1E)。KEGG結(jié)果顯示RMRP在酒精中毒、系統(tǒng)性紅斑狼瘡、病毒癌變、轉(zhuǎn)錄異常等途徑富集(圖1F)。然后進(jìn)行KEGG研究的基因集富集分析(GSEA)。RMRP在細(xì)胞周期、結(jié)直腸癌、蛋白質(zhì)輸出、TGFB途徑和泛素介導(dǎo)的蛋白水解途徑中富集(圖1G)。這些通路與腫瘤的發(fā)生和轉(zhuǎn)移密切相關(guān),提示RMRP可能在NSCLC中發(fā)揮重要作用。我們還對RMRP的蛋白編碼潛能進(jìn)行了探究,發(fā)現(xiàn)RMRP不具有蛋白編碼潛能(圖1H和I)。
2)m6A修飾在RMRP中富集,提高了RMRP的轉(zhuǎn)錄穩(wěn)定性
最近的初步研究報(bào)道,m6A修飾廣泛存在,通過調(diào)控轉(zhuǎn)錄組影響RNA的剪接、翻譯、輸出、定位和穩(wěn)定性。為了探究m6A對RMRP的修飾,我們首先使用在線生物信息學(xué)工具m6Avar預(yù)測位于RMRP中的m6A位點(diǎn),并鑒定出兩個(gè)RMRP m6A序列基元。然后,在人肺上皮細(xì)胞(HBE)和兩株NSCLC細(xì)胞株(A549和H1299)中進(jìn)行甲基化RNA免疫沉淀(Me-RIP)檢測。MeRIP-qPCR檢測顯示,HBE細(xì)胞中RMRP的m6A甲基化水平低于NSCLC細(xì)胞(A549和H1299)(圖2A)。然后我們用小干擾RNA靶向m6A甲基化酶復(fù)合物的核心成分METTTL3,發(fā)現(xiàn)A549細(xì)胞中總RNA和RMRP RNA中的m6A水平都降低了(圖2B, C, D)。放線菌素D 阻斷A549細(xì)胞新RNA合成后,檢測RMRP RNA的丟失情況。結(jié)果顯示,METTL3下調(diào)后,RMRP表現(xiàn)出較低的RNA穩(wěn)定性(圖2E)。

3)RMRP促進(jìn)NSCLC細(xì)胞增殖
為了進(jìn)一步探討RMRP的作用,我們采用定量RT-PCR方法檢測RMRP在NSCLC細(xì)胞中的表達(dá)。RMRP在A549細(xì)胞中的表達(dá)明顯高于H1299細(xì)胞(圖3A)。因此,我們在A549細(xì)胞中轉(zhuǎn)染了兩種不同的抗RMRP shRNA (shRMRP-1和shRMRP-2)或?qū)φ战M的shRNA (rash),并在H1299細(xì)胞中轉(zhuǎn)染了RMRP過表達(dá)質(zhì)粒(RMRP)或?qū)φ盏馁|(zhì)粒。通過定量RT-PCR分析證實(shí)了轉(zhuǎn)染效率(圖3B和C)。CCK-8和菌落形成實(shí)驗(yàn)表明,轉(zhuǎn)染了shRMRP的NSCLC細(xì)胞的生長和增殖受到了顯著的抑制。而轉(zhuǎn)染過表達(dá)RMRP的質(zhì)粒后效果相反(圖3D-G)。

4)TGFBR1是RMRP促進(jìn)增殖、侵襲和遷移的關(guān)鍵靶點(diǎn)
在我們之前的研究中,我們發(fā)現(xiàn)TGFBR1與NSCLC的增殖和轉(zhuǎn)移相關(guān),GSEA結(jié)果顯示RMRP可能參與了TGFB通路。TCGA數(shù)據(jù)庫顯示TGFBR1與RMRP表達(dá)呈正相關(guān)(圖4A)。因此,我們假設(shè)RMRP可能以TGFBR1為靶點(diǎn)。此外,在A549或H1299細(xì)胞中,RMRP敲低或過表達(dá)后,TGFBR1的表達(dá)發(fā)生改變(圖4B、C、D)。在A549細(xì)胞中,下調(diào)RMRP后,TGFBR1的表達(dá)降低,BAX/ Bcl-2升高,而TGFBR1過表達(dá)后,效果相反。在H1299細(xì)胞中,過表達(dá)RMRP后,TGFBR1的表達(dá)增加,BAX/Bcl-2的表達(dá)減少,而TGFBR1敲低后則相反(圖4D)。此外,我們還探索了TGFBR1參與RMRP促進(jìn)NSCLC生長和增殖的機(jī)制。結(jié)果顯示,TGFBR1過表達(dá)后,shRMRP對NSCLC細(xì)胞生長和增殖的抑制作用得到恢復(fù),而TGFBR1敲低后則相反(圖4E, F, H, J)。
TGFBR1促進(jìn)NSCLC細(xì)胞的侵襲、遷移和G1細(xì)胞周期進(jìn)展。因此,我們探討了RMRP對侵襲、遷移和G1細(xì)胞周期進(jìn)程的影響。與對照組相比,轉(zhuǎn)染shRMRP的NSCLC細(xì)胞顯著抑制了侵襲、遷移和G1細(xì)胞周期進(jìn)展,而在RMRP過表達(dá)的細(xì)胞中觀察到相反的效果。在TGFBR1過表達(dá)后,,shRMRP對NSCLC細(xì)胞的侵襲、遷移和G1細(xì)胞周期進(jìn)展的抑制作用被挽救,而在TGFBR1敲低后,觀察到相反的效果(圖4G, I, K, L, M,和N)。以上結(jié)果表明TGFBR1可能是RMRP促進(jìn)增殖、侵襲和遷移的關(guān)鍵靶點(diǎn)。

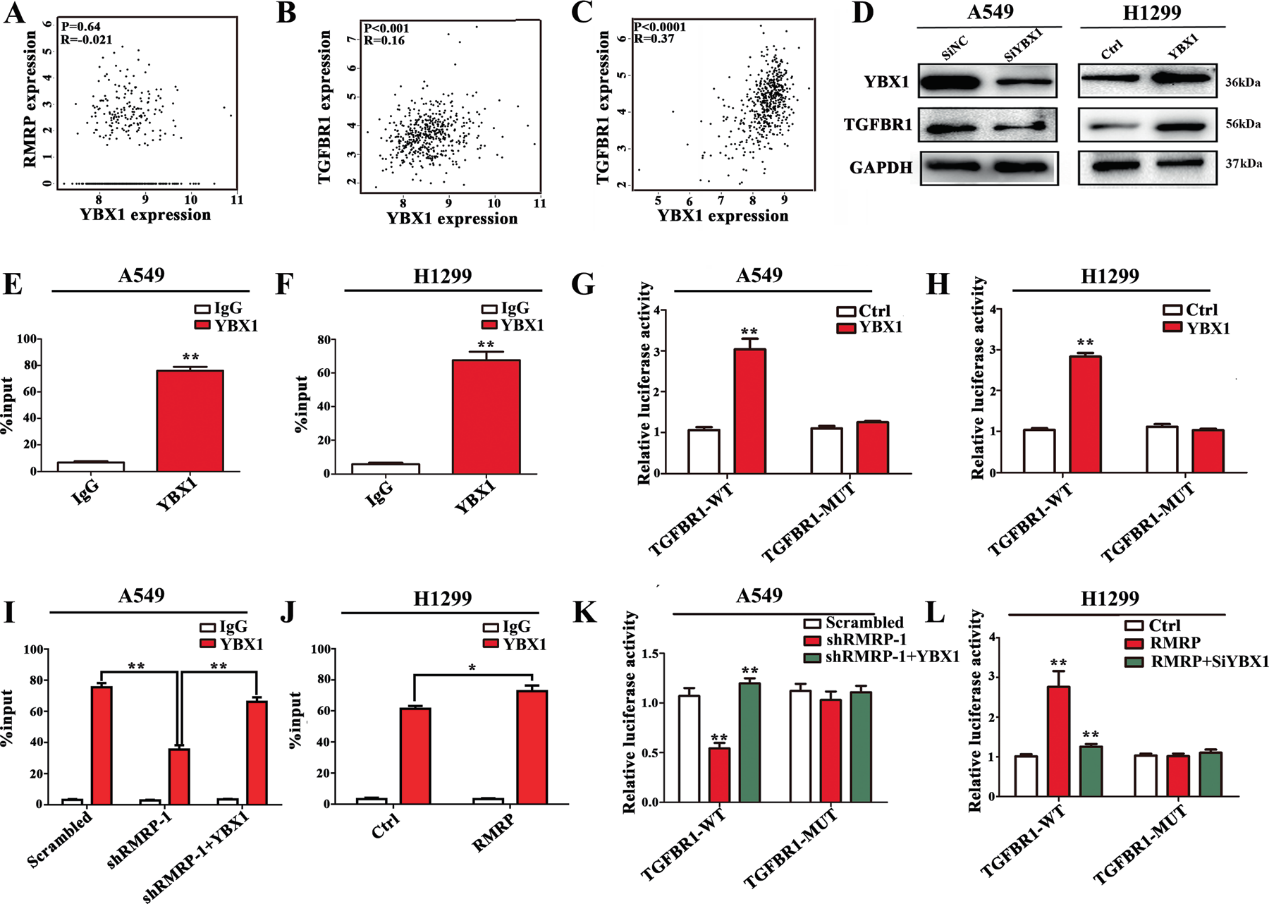
5)RMRP與轉(zhuǎn)錄因子YBX1相關(guān)
為了進(jìn)一步闡明RMRP的機(jī)制,我們在NSCLC細(xì)胞中進(jìn)行了核和細(xì)胞質(zhì)RNA的分離和熒光原位雜交(FISH)檢測。結(jié)果顯示,RMRP主要位于胞質(zhì)(圖5A和B)。最近的一些報(bào)道發(fā)現(xiàn),許多l(xiāng)ncRNA通過與DNA或蛋白質(zhì)相互作用發(fā)揮作用。為了探討RMRP的調(diào)控作用,我們使用catRAPID。結(jié)果顯示,在NSCLC中發(fā)揮重要作用的YBX1是RMRP的潛在結(jié)合蛋白(圖5C)。RIP實(shí)驗(yàn)顯示,抗YBX1抗體顯著富集了RMRP(圖5D和E)。RNA下拉實(shí)驗(yàn)進(jìn)一步證實(shí)了RMRP和YBX1之間的相互作用(圖5F)。以上結(jié)果表明YBX1與RMRP之間存在密切的相互作用。

6)RMRP通過招募YBX1來促進(jìn)TGFBR1的轉(zhuǎn)錄
我們探索TCGA-LUAD的ATAC-seq數(shù)據(jù),以確定TGFBR1可能的轉(zhuǎn)錄因子。該分析揭示了TGFBR1基因啟動(dòng)子區(qū)可能存在的YBX1結(jié)合序列。在GEPIA的TCGA-LUAD數(shù)據(jù)庫中,YBX1的表達(dá)與TGFBR1呈正相關(guān),而與RMRP無相關(guān)性(圖6A-C)。因此,我們推測YBX1是一個(gè)轉(zhuǎn)錄因子,可能被RMRP招募到TGFBR1啟動(dòng)子中。為了進(jìn)一步研究其機(jī)制,我們分別轉(zhuǎn)染siRNA (SiYBX1)、pcDNAYBX1 (YBX1)和它們匹配的對照(SiNC或Ctrl)。YBX1敲低抑制了TGFBR1的表達(dá),而YBX1過表達(dá)則增加了TGFBR1的表達(dá)(圖6D)。YBX1結(jié)合復(fù)合物顯示TGFBR1啟動(dòng)子顯著富集。YBX1過表達(dá)增加了TGFBR1-WT細(xì)胞中的熒光素酶活性 (圖6G和H)。RMRP敲低降低了YBX1在TGFBR1啟動(dòng)子上的富集。共轉(zhuǎn)染YBX1可逆轉(zhuǎn)這一效應(yīng)(圖6I)。此外,RMRP增加了結(jié)合(圖6J)。熒光素酶檢測進(jìn)一步顯示,熒光素酶活性的變化是由于RMRP的敲減或過表達(dá)(圖6K和L)。
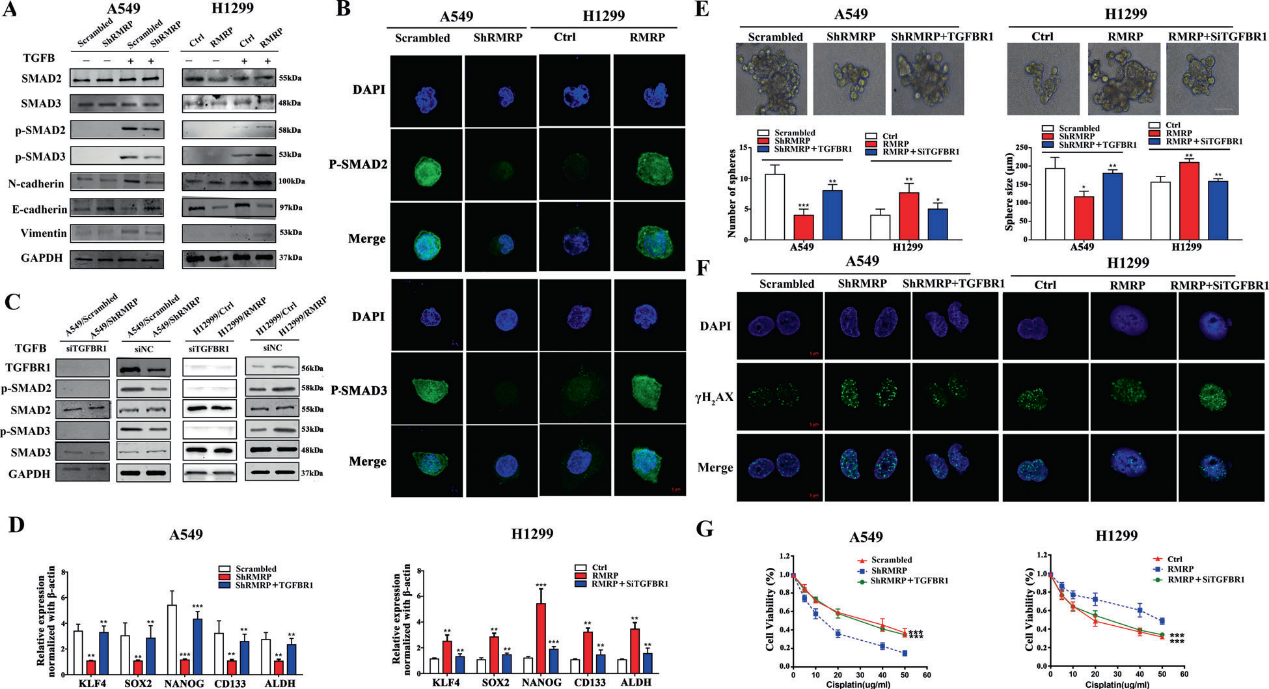
7)RMRP調(diào)控NSCLC中TGFBR1/SMAD2/ SMAD3通路
TGFBR1/SMAD2/SMAD3通路在NSCLC中發(fā)揮重要作用。Western blot和免疫熒光檢測顯示,RMRP敲低顯著抑制了NSCLC細(xì)胞中的核易位和p-SMAD2/ 3的表達(dá)(圖7A和B)。RMRP的改變對siTGFBR1組中p-SMAD2和p-SMAD3的表達(dá)水平?jīng)]有影響,這表明TGFBR1是RMRP激活SMAD2/SMAD3信號通路所必需的(圖7C)。
TGFB通路常與CSC性質(zhì)和EMT有關(guān)。因此,我們探討了RMRP是否促進(jìn)NSCLC中的CSC特性。我們通過qRT-PCR檢測了CSC相關(guān)基因KLF4、SOX2、NANOG、CD133和ALDH的表達(dá),發(fā)現(xiàn)在A549細(xì)胞中,RMRP敲低后這些基因的表達(dá)降低。在H1299細(xì)胞中,過表達(dá)RMRP后,這些基因表達(dá)上調(diào)(圖7D)。此外,我們還進(jìn)一步探索了這些細(xì)胞中EMT相關(guān)基因的表達(dá)。結(jié)果顯示,RMRP敲低增加了E-Cadherin的表達(dá),降低了N-Cadherin和Vimentin的表達(dá)(圖7A)。我們還探索了這些細(xì)胞的成球能力。實(shí)驗(yàn)表明,RMRP敲低降低了成球能力,而過表達(dá)RMRP則增加了成球能力(圖7E)。RMRP抑制增加了A549細(xì)胞對順鉑和放療的敏感性(圖7F和G)。這些數(shù)據(jù)表明,RMRP通過增強(qiáng)CSC自我更新和EMT促進(jìn)NSCLC進(jìn)展。
8)在體內(nèi)抑制RMRP抑制腫瘤生長
為了進(jìn)一步證實(shí)RMRP在體內(nèi)促進(jìn)腫瘤生長,將A549-shRMRP細(xì)胞和A549- scramble細(xì)胞注射到裸鼠體內(nèi)。30天后,shRMRP組腫瘤明顯變小(圖8A和B)。shRMRP組腫瘤體積和重量顯著降低(圖8C,D)。此外,在A549-scramble中,Ki-67、MMP9和TGFBR1的表達(dá)更高(圖8E)。

結(jié)論:m6A修飾促進(jìn)了lncRNA-RMRP/TGFBR1/SMAD2/SMAD3通路。其潛在的機(jī)制是通過RMRP將轉(zhuǎn)錄因子YBX1招募到TGFBR1啟動(dòng)子,從而對TGFBR1進(jìn)行轉(zhuǎn)錄調(diào)控。此外,RMRP增加了球形成能力和EMT,這與放療和化療的耐藥有關(guān)。RMRP是一種生物標(biāo)志物,在NSCLC中具有潛在的預(yù)后和治療相關(guān)性。
參考文獻(xiàn):Yin H, Chen L, Piao S, Wang Y, Li Z, Lin Y, Tang X, Zhang H, Zhang H, Wang X. M6A RNA methylation-mediated RMRP stability renders proliferation and progression of non-small cell lung cancer through regulating TGFBR1/SMAD2/SMAD3 pathway. Cell Death Differ. 2021 Oct 9. doi: 10.1038/s41418-021-00888-8. Epub ahead of print. PMID: 34628486.