






RAS GTPase RIT1通過抑制紡錘體組裝檢查點降低有絲分裂的保真度

2021年6月,最新一篇發(fā)表在Curr Biol(IF=9.601)的文章(The RAS GTPase RIT1 compromises mitotic fidelity through spindle assembly checkpoint suppression.)從共生功能體視點的角度對29名接受同種異體造血干細胞移植的兒童的腸道、口腔和鼻腔微生物群進行了綜合宿主微生物群分析。首次揭示了RIT1直接與SAC核心組件MAD2和p31conmet相互作用;CDK1在有絲分裂過程中磷酸化RIT1并抑制其與SAC的相互作用;RIT1是細胞有絲分裂及時進展所必需的;RIT1致病水平促進染色體分離錯誤。
背景:RIT1突變已被確定為肺腺癌的致癌因素和努南綜合征的病因因素。RIT1有一組獨特的效應蛋白,但與其他Ras GTPases共享MAPK通路的激活。然而,由于缺乏確定的同源GTPase激活蛋白或交換因子,RIT1 GTPase周期的調控仍不清楚。盡管如此,RIT1的豐度和活性是通過蛋白酶體降解在蛋白水平上調控的,這種機制是由適配器蛋白LZTR1和E3泛素連接酶Cullin 3 (CRL3LZTR1)介導的。RIT1在Noonan綜合征中的作用很可能是通過MAPK通路的過度激活介導的,MAPK通路是該疾病的一種癥狀體征,但它在正常細胞和惡性腫瘤中的作用尚不清楚.
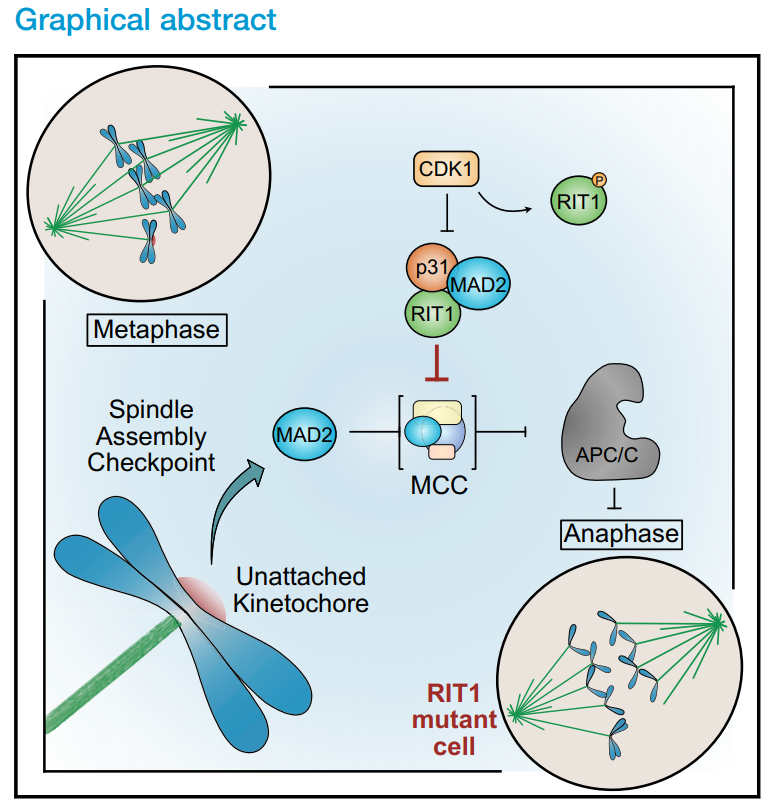
紡錘體裝配檢查點(SAC)作為一個傳感器的獨立的著絲點延遲有絲分裂進入后期,直到適當?shù)娜旧w分離得到保證。這一安全機制的破壞導致基因組不穩(wěn)定和非整倍體,這是胚胎死亡、先天性出生缺陷、智力殘疾和癌癥的遺傳原因。然而,盡管了解了控制SAC的基本機制,但信號通路如何直接與有絲分裂檢查點活動相互作用和調節(jié)仍是未知的。在對細胞外刺激的反應中,參與細胞生長、生存和分化的多種信號通路網(wǎng)絡被激活,這一過程顯著受到Ras家族小鳥苷三磷酸酶(GTPases)的調控。RIT1是一種ras相關的GTPase,調節(jié)細胞存活和應激反應,是通過有絲分裂和適當?shù)娜旧w分離及時進展的必要條件。RIT1在有絲分裂期間從質膜(PM)分離,并直接與SAC蛋白MAD2和p31conmet相互作用,該過程受周期蛋白依賴性激酶1 (CDK1)活性的調節(jié)。此外,致病水平的RIT1沉默了SAC,并通過將MAD2從有絲分裂檢查點復合體(MCC)中分離出來,加速了通過有絲分裂的轉運。此外,致病性RIT1抑制SAC促進染色體分離錯誤和非整倍體。我們的研究結果突出了RIT1與其他Ras GTPases相比的獨特功能,并闡明了信號通路與SAC之間通過一種新的調節(jié)機制的直接聯(lián)系。
一、RIT1直接與SAC蛋白MAD2和p31conmet相互作用

為了表征RIT1相互作用組,我們進行了親和純化-質譜篩選(圖1A),確定MAD2 (MAD2L1)和p31conmet(也稱為MAD2L1結合蛋白)作為新的和選擇性的RIT1結合伙伴,不與其他Ras GTPases相互作用(圖S1A和S1B)。
MAD2參與了在未附著的動點處催化MCC形成的SAC信號放大,MCC由MAD2、CDC20、BubR1和Bub3.1組成。16 MAD2和p31conmet二聚促使我們評估RIT1是否與MAD2和p31conmet直接相互作用。Pull down分析顯示,這兩種相互作用都直接且獨立于MAD2和p31conmet二聚(圖1B)。此外,RIT1-MAD2相互作用在斑馬魚和果蠅中是保守的(圖1C)。為了確定RIT1與MAD2或p31conmet的結合是否受其GTPase周期的調控,我們評估了與GTPgS(一種不可水解的GTP類似物)加載的RIT1的結合。這表明這兩種相互作用都不依賴于RIT1的鳥苷核苷酸負載狀態(tài)(圖1D)。一致地,與MAD2和p31conmet的結合不受與疾病相關的RIT1突變的影響(圖S1C)。這些結果表明,結合界面位于對GDP/GTP結合敏感的RIT1 s開關I和II結構域外,因此MAD2和p31conmet不是典型的RIT1效應蛋白。MAD2和p31conmet的結構相似性突出了RIT1的潛在結合競爭。因此,我們使用了競爭結合試驗,其中滴定MAD2野生型(WT)或R133E/Q134A (RQ),一個二聚體和p31結合缺陷的突變體,未能抑制rit1 -p31conmet結合(圖1E)。由于RIT1 g結構域與其他Ras - GTPases,特別是它的鄰域RIT2之間的相似性,假設RIT1 s的n端或c端延伸可能介導與MAD2和p31conmet的相互作用(圖1F)。隨后分析了RIT1 N-或c -末端缺失突變體,證明了c -末端結構域對于MAD2和p31conmet星結合是必要和充分的(圖1G、1H和S1J)。
二、RIT1與MAD2和p31conmet的相互作用由CDK1磷酸化調控
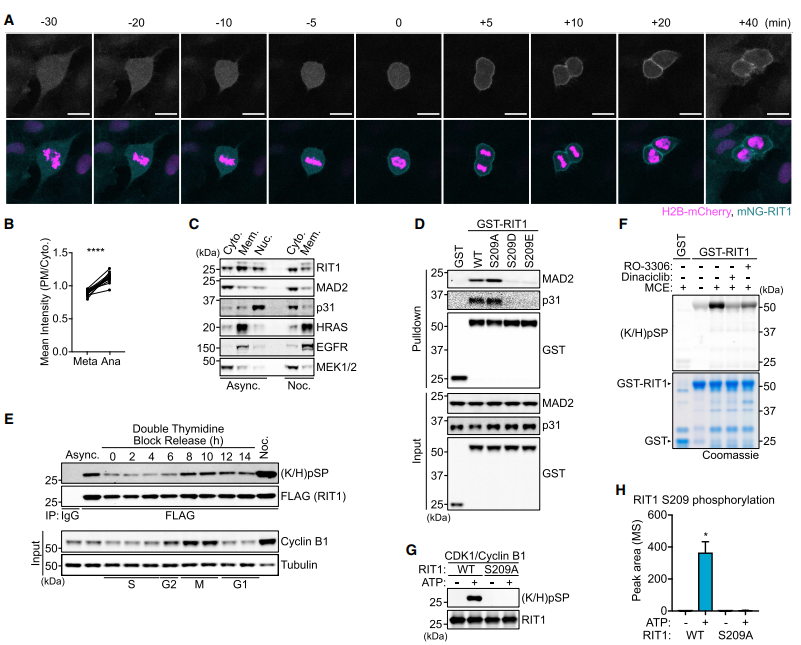
在間期,RIT1 s c端尾部介導質膜(PM)結合然而,在分析有絲分裂細胞時,我們觀察到RIT1在細胞進入有絲分裂和進入中期并在后期快速易位到PM時的彌漫性胞質分布(圖2A和2B)。同樣,在有絲分裂細胞裂解物中檢測到內源性RIT1的主要胞質分布(圖2C)。(S209D/E)磷酸化,而非(S209A)缺磷,突變破壞了RIT1-MAD2/p31conmet結合(圖2D)。RIT1磷酸化在有絲分裂過程中最為豐富(圖2E)。抑制CDK1顯著降低了重組RIT1的磷酸化(圖2F)。免疫印跡檢測和質譜證實CDK1/CyclinB1磷酸化RIT1 S209,(圖2G和2H)。
三、RIT1及時調節(jié)后期進入和染色體分離保真度

MAD2和p31conmet調節(jié)SAC的持續(xù)時間,反過來,也調節(jié)有絲分裂的持續(xù)時間。這促使我們研究RIT1是否通過與MAD2和p31conmet的直接關聯(lián)來影響SAC。通過RNAi或crispr介導的敲除去除RIT1可延長有絲分裂進程(圖3A和S3A S3E)。此外,SAC的藥理抑制挽救了RIT1耗盡的作用,表明RIT1以SAC依賴的方式影響有絲分裂。此外,RIT1的缺失增加了染色體分離錯誤的發(fā)生率(圖3B),這表明RIT1不僅對有絲分裂的及時進展至關重要,而且RIT1蛋白水平的失調也破壞了正常的SAC功能。LZTR1或RIT1M90I表達的缺失加速了異步生長細胞的有絲分裂進程,這一效應依賴于PM釋放RIT1(圖3C、S3H和S3I)。同樣,RIT1 WT或M90I的過表達部分覆蓋了藥物誘導的SAC反應(圖3D和S3J)。RIT1M90I的異位表達以依賴MAD2-和p31come結合的方式顯著增加了有絲分裂錯誤的發(fā)生率,包括滯后染色體和橋接染色體(圖3G和S3O)。因此,我們觀察到在表達RIT1M90I的細胞中非整倍體率增加,但在表達不能結合MAD2/p31conmet的突變體的細胞中卻沒有(圖3H和S3P)。這些結果表明,RIT1水平的增加會導致與MAD2和p31conmet的直接相互作用,從而降低有絲分裂的保真度。
四、RIT1抑制MCC-MAD2結合,促進APC/C底物降解
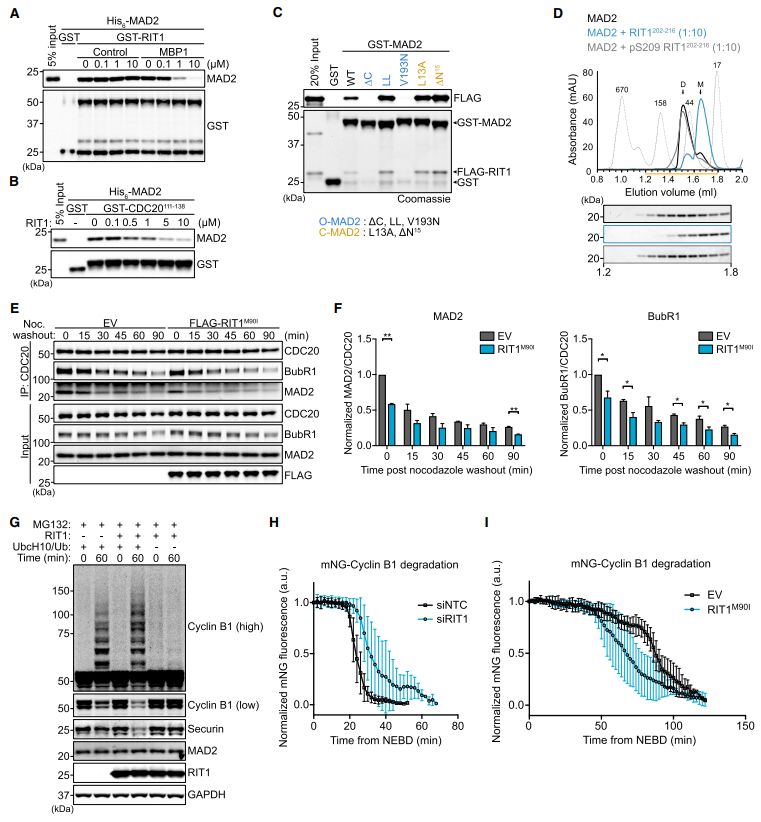
在MAD1與未連接的動點結合后,MAD2從開放的構象狀態(tài)(O-MAD2)轉化為封閉的構象狀態(tài)(C-MAD2), SAC信號被緊密調控和放大。MAD2的構象變化啟動了它與CDC20的結合。為了直接測試RIT1是否抑制了MAD2與CDC20或MAD1的關聯(lián),進行了競爭性pull-down實驗來測試RIT1-MAD2和MAD2-CDC20/ MAD1結合之間的互斥性(圖4A和4B)。MAD2結合肽1 (MBP1)是一種模擬CDC20和MAD1相互作用基序(MIM)的高親和合成肽,它取消了MAD2- rit1的結合。評估了RIT1與O-或c -狀態(tài)穩(wěn)定的MAD2突變體的結合(圖4C)。用過量的RIT1 c端尾肽孵育MAD2蛋白,并用凝膠過濾分離(圖4D)。RIT1M90I顯著降低了MAD2- CDC20和BubR1-CDC20的相互作用(圖4E和4F);表明致病RIT1蛋白水平阻礙MCC的完整性,符合RIT1從CDC20中隔離MAD2并促進MCC拆卸的模型。
用重組RIT1補充這些提取物增加了CyclinB1和Securin的泛素化和降解,表明APC/C活性增加,可能是由于MCC抑制緩解(圖4G)。RIT1缺失細胞顯示CyclinB1降解延遲(圖4H)。此外,RIT1M90I的表達加速了正常細胞生長下CyclinB1的降解(圖4I);然而,其對CyclinB1降解的影響在藥理學誘導的有絲分裂阻滯下被消除(圖S4M),這表明致病性水平的RIT1不能抑制過度活躍的SAC反應。