lncRNA INCR1調節(jié)IFNγ信號逃避腫瘤免疫
導讀:
免疫檢查點抑制劑治療是基于腫瘤上調免疫檢查點分子(如PD-L1)來逃避抗腫瘤免疫的能力,然而部分應答者對這些療法出現(xiàn)獲得性耐藥。免疫檢查點抑制劑的耐藥部分是由于腫瘤中IFN刺激基因的持續(xù)表達,導致持續(xù)的IFNγ信號傳導。lncRNA轉錄過程本身可以促進染色質重塑,增強編碼基因啟動子的活性。研究顯示lncRNA的異常表達與腫瘤的發(fā)生發(fā)展有關,但它們在免疫逃避中的作用仍不清楚。Mineo等人在下面這篇文章中研究了lncRNA INCR1在腫瘤免疫逃避中的作用。

技術路線圖:
結果:
1.lncRNA INCR1在IFNγ刺激的腫瘤細胞中表達
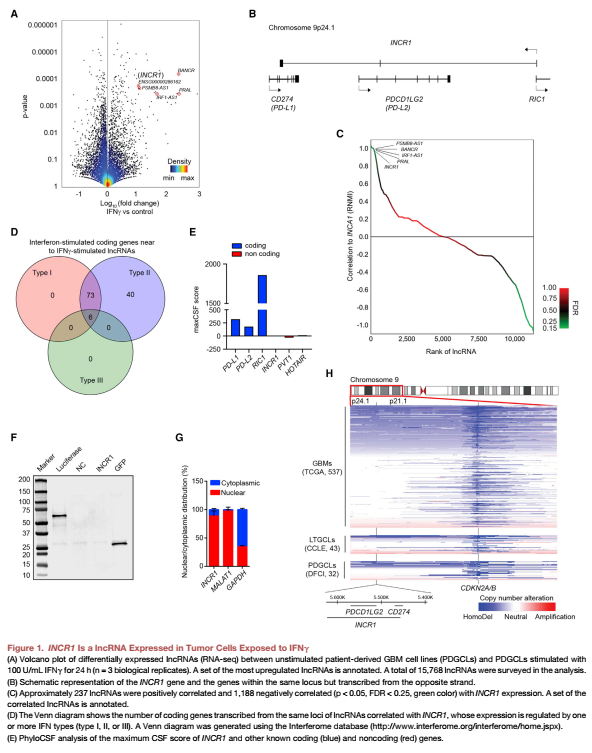
首先對IFNγ刺激的患者源性GBM細胞株(PDGCLs)進行了全轉錄組分析(RNA測序,F(xiàn)igure 1A),IFNγ刺激了113個lncRNA的轉錄,包括已經被驗證的IFNγ刺激之后上調的lncRNA BANCR。在上調最顯著的lncRNA中,有一個表達于PD-L1/PD-L2位點DNA鏈的lncRNA特征不明顯,命名為lncRNA INCR1(Figure 1B)。INCR1表達與237個lncRNAs表達呈正相關( Figure 1C),其中幾個轉錄于受IFN調控的編碼蛋白質的基因座(Figure 1D)。PhyloCSF對INCR1序列進行分析,結果顯示該轉錄本具有低編碼潛力( (Figure 1E))。通過體外轉錄/翻譯實驗,文章也證實了INCR1沒有被翻譯成蛋白( (Figure 1F)。qPCR分析顯示,INCR1主要定位在細胞核內(Figure 1G)。分析INCR1位點和附近的9p21細胞帶,包括CDKN2A/B基因,在32個PDGCLs和43個長期GBM細胞系(LTGCLs)中,顯示了與來自癌癥基因組圖譜(TCGA)隊列的GBM腫瘤相似的拷貝數(shù)改變模式,這表明實驗中的細胞系模型反映了人類疾病。
2.在不同的癌細胞和病人的腫瘤中,INCR1表達與PD-L1水平相關
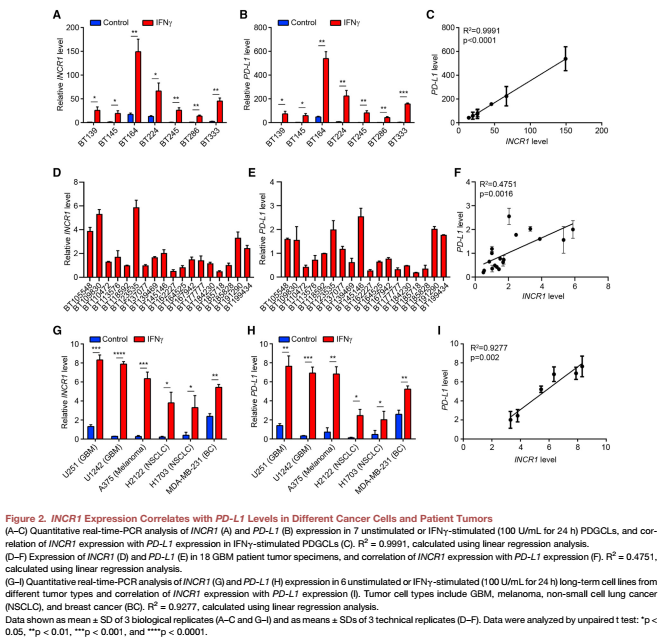
對PDGCLs 進行qPCR分析發(fā)現(xiàn)在所有的細胞系中,IFNγ刺激INCR1上調 (Figure 2A)。IFNγ處理的PDGCLs也表達高水平的PD-L1 mRNA和蛋白(Figure 2B)。值得注意的是,我們觀察到INCR1和PD-L1 mRNA和蛋白之間高度顯著的相關性(Figure 2C)。與這些發(fā)現(xiàn)相一致的是,INCR1在患者GBMs中表達與PD-L1 mRNA水平呈正相關,INCR1高的腫瘤也表達較高的PD-L1水平(Figure 2D and 2F)。實驗選擇6個代表GBM、黑色素瘤、NSCLC和BC的細胞系。IFNγ刺激后,所有細胞系均顯示INCR1表達增加(Figure 2G),這也與PD-L1表達相關(Figures 2H and 2I). 因此,說明文章發(fā)現(xiàn)了腫瘤和組織培養(yǎng)細胞中一種新的lncRNA,IFN刺激其表達,其表達與PD-L1呈正相關。
3.INCR1調節(jié)腫瘤IFNγ信號通路
作者推測INCR1會調節(jié)其鄰近基因的表達和IFNγ刺激引起的腫瘤應答。沉默INCR1降低了938個基因的表達,其中451個基因在未受刺激和IFNγ刺激的細胞中普遍下調(Figure 3A )。基因本體論(GO)富集分析顯示下調的基因參與免疫相關功能,如IFNγ應答、先天免疫應答和對病毒的防御應答(Figure 3B)。此外,INCR1基因敲低導致124個ISGs基因表達減少(Figure 3C)。這些基因中包括IFNγ信號通路的重要組成部分(JAK2,STAT1),以及主要的免疫抑制分子(PD-L1和IDO1)。研究證實,無論是在基礎水平上還是在IFNγ作用下,沉默INCR1都會導致其重疊基因PD-L1和PDL2的mRNA水平降低(Figure 3D 3F)。值得注意的是,與INCR1和PD-L1在同一位點的JAK2的mRNA顯著下調(Figure 3G),來自不同基因組區(qū)域的ISGs如STAT1和IDO1的mRNA也顯著下調(Figure 3H和3I)。此外,文章中證明了沉默INCR1導致PD-L1、JAK2、STAT1和IDO1蛋白水平下調(Figure 3J)。總STAT1的下調與IFNγ刺激下其磷酸化和核定位降低相關(Figure 3J)。此外,我們發(fā)現(xiàn)沉默INCR1降低了PD-L1總蛋白和細胞表面PD-L1水平。
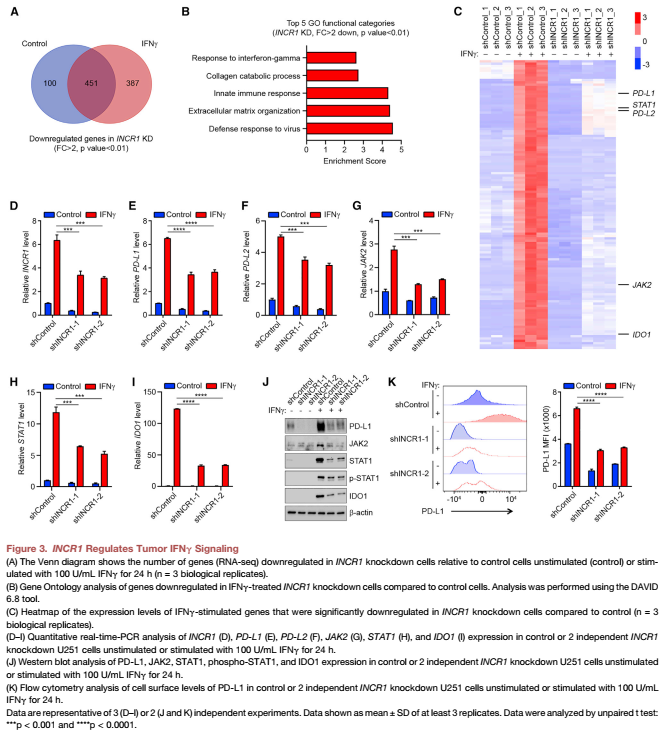
4.體外沉默INCR1導致T細胞介導的細胞毒性增加;體內提高CAR-T細胞療效
研究表明CD8+ CTLs的激活誘導IFNγ等細胞因子的分泌,促進其增殖和抗腫瘤活性。將腫瘤細胞與CD8 + CTL共培養(yǎng),用antiCD3和anti-CD28抗體共價偶聯(lián)的珠子激活,會使INCR1沉默細胞的細胞毒性比對照組增加,但活化T細胞的生存能力沒有顯著變化(Figure 4A)。使用3D培養(yǎng)系統(tǒng),我們發(fā)現(xiàn)當INCR1敲除腫瘤球與激活的CD8+ ctl共孵養(yǎng)時顯著減小,與對照腫瘤球相比(Figure 4B)。這與INCR1沉默細胞共培養(yǎng)的CD8+ CTLs中IFNγ分泌增加有關,與CD8+ CTLs共培養(yǎng)的空白細胞相比。
將U251-EGFRvIII對照和INCR1沉默腫瘤植入皮下。植入后7天,單獨表達GFP的人T細胞(對照組)或EGFRvIII-directed CAR被單次靜脈注射為標準劑量的1/10。在這些條件下,對照組腫瘤小鼠對CAR-T細胞治療沒有明顯反應。相比之下,CAR-T細胞顯著降低了INCR1敲低腫瘤小鼠的腫瘤生長。在研究結束時,也就是注射T細胞后的21天,我們對腫瘤進行了CAR - T細胞存在的分析。INCR1敲除腫瘤同時出現(xiàn)CD4+和CD8+ T細胞浸潤,以CD8+ T細胞為主。相比之下,對照組腫瘤未見CD4+ T細胞浸潤(Figure 4E,4F)。此外,與INCR1沉默的浸潤性腫瘤相比,對照組的CD8+ T細胞表達的PD-1水平明顯更高(Figure 4G)。這些結果表明INCR1在控制腫瘤IFNγ信號中發(fā)揮著功能作用,其敲除導致人類腫瘤細胞對T細胞介導的殺傷的敏感性增加。
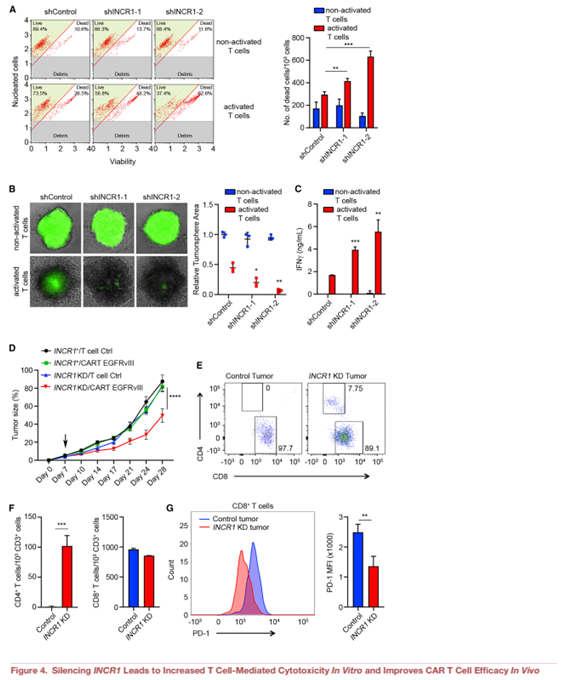
5.HNRNPH1是INCR1的結合搭檔
大多數(shù)lncrna通過與蛋白質相互作用而發(fā)揮作用。為了明確INCR1調控PD-L1表達的機制,我們采用RNA反義純化(RAP)對IFNγ刺激的PDGCLs中與內源性INCR1直接接觸的蛋白進行純化。我們設計了覆蓋整個INCR1序列的探針,通過UV照射交聯(lián)RNA -蛋白復合物,并在變性條件下進行RAP,以最大限度地恢復特定RNA-蛋白相互作用。與對照純化相比,我們觀察到大于80倍的INCR1 RNA富集((Figure 5A)。質譜分析顯示,在INCR1 RAP中,HNRNPH1蛋白結合最高,而在對照RAP中則不是(Figure 5B)。然后,我們通過體內RNA UV交聯(lián)和免疫沉淀(CLIP)驗證了INCR1和HNRNPH1之間的相互作用。此外,與未處理的細胞和同型對照相比,IFNγ刺激的細胞中HNRNPH1與INCR1顯著結合(Figure 5C)。為了識別HNRNPH1結合的INCR1區(qū)域,我們首先通過增強剪輯測序(eclipseq)繪制了體內HNRNPH1結合位點。我們在INCR1的近端內含子上發(fā)現(xiàn)了一簇峰 (Figure 5D)。
為了在體外驗證測序數(shù)據(jù),我們克隆了一個包含第一個內含子50和30區(qū)域的INCR1小基因,并利用它生成了7個不同的體外轉錄的生物素化RNA。RNA pull-down分析顯示,HNRNPH1與包含在eclipseq中發(fā)現(xiàn)的主要峰序列的2個RNA片段(F4和F6)相互作用強烈(Figure 5E)。HNRNPH1結合的兩個片段在G-stretches中富集,該基序在80%的HNRNPH1 RNA靶標中被發(fā)現(xiàn)(Figure5F)。GO富集分析所有被eCLIP-seq確定的基因,顯示HNRNPH1,除了結合到INCR1,也結合到參與免疫系統(tǒng)過程的多個基因,包括幾個IFNγ刺激的基因如PD-L1 JAK2和STAT1,它們的表達受INCR1調節(jié)(Figure 5G)。接下來,我們通過RNA免疫沉淀(RIP)驗證HNRNPH1結合PD-L1和JAK2 mRNA(Figure 5H 5I)。這些結果表明INCR1與HNRNPH1的結合不依賴于PD-L1和JAK2 mRNA的結合。
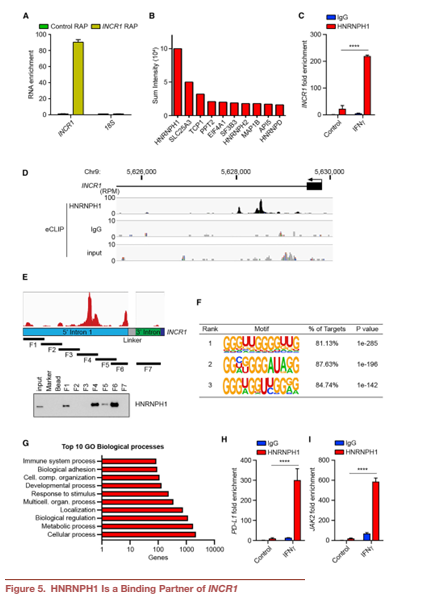
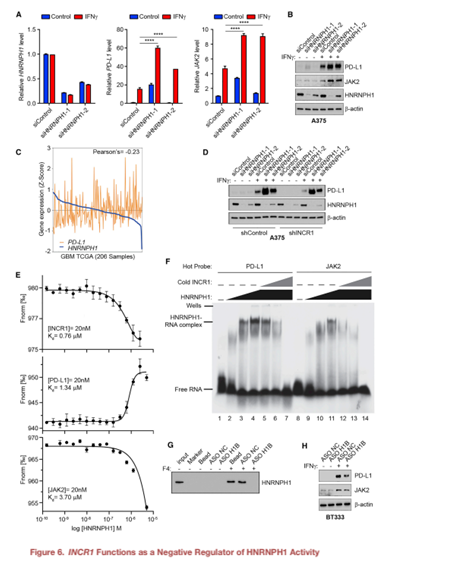
6. INCR1是HNRNPH1活性的負調控因子
沉默HNRNPH1可以提高IFNγ處理的A375細胞((Figure 6A)和患者源性BT139細胞的PD-L1和JAK2 mRNA水平。這與IFNγ刺激下PD-L1和JAK2蛋白顯著增加有關(Figure 6B)。這些數(shù)據(jù)提示HNRNPH1是PD-L1和JAK2表達的負調控因子,需要結合INCR1才能阻止HNRNPH1的功能。在GBM腫瘤(TCGA)中,觀察到HNRNPH1水平與PD-L1表達呈負相關,支持了這一假設(Figure 6C)。為了進一步證明我們的假設,我們進行了敲低實驗來沉默INCR1敲低細胞中HNRNPH1的表達。正如預期的那樣,INCR1敲低導致IFNγ處理的細胞中PD-L1表達減少。然而,沉默敲除細胞中的HNRNPH1挽救了PD-L1的表達(Figure 6D)。與PD-L1和JAK2 RNA片段相比,INCR1 RNA片段對HNRNPH1表現(xiàn)出更高的結合親和力(Figure 6E)。這些數(shù)據(jù)表明,HNRNPH1與INCR1結合可能會降低其與PD-L1和JAK2相互作用的能力。利用g-32P放射性標記的PD-L1和JAK2 RNA片段,我們觀察到RNA:HNRNPH1復合物的位移,證實了這兩個序列結合HNRNPH1的能力(Figure 6F)。INCR1:PD-L1和INCR1:JAK2比值為1:1中非標記INCR1 RNA片段的添加有效降低PD-L1:HNRNPH1和JAK2:HNRNPH1復合物的形成(Figure 6F),表明HNRNPH1與JAK2之間的親和力弱于HNRNPH1與PD-L1之間的親和力。將摩爾比增加到1:5和1:10,復合物完全解離(Figure 6F)。這些結果表明INCR1對HNRNPH1的親和力強于對PD-L1或JAK2,并且INCR1抑制了HNRNPH1與PDL1和JAK2的相互作用。
通過ASO H1B(靶向INCR1 RNA中HNRNPH1結合位點),我們可以在體外以劑量依賴的方式減少INCR1與HNRNPH1的相互作用(Figure 6G)。然而,使用對照ASO檢測未觀察到對結合能力的影響(Figure 6G)。為了研究ASO H1B在體內的作用,在ASO轉染BT333患者源性細胞和A375黑色素瘤細胞中,ASO H1B均顯著降低IFNγ刺激的PD-L1和JAK2蛋白的表達(Figure 6H)。這些結果表明INCR1特異性地與HNRNPH1相互作用,阻斷這種相互作用會影響PD-L1和JAK2的表達。