






人18S rRNA m6A甲基轉移酶新發現——METTL5
RNA甲基化在mRNA上被大量發現,rRNA甲基化修飾卻研究的很少,并且rRNA甲基化修飾作用仍不清楚。近日,來自法國巴黎理工學院的研究團隊在Nucleic Acids Research期刊上發表題名為:The human 18S rRNA m6A methyltransferase METTL5 is stabilized by TRMT112的研究文章。該文章主要發現METTL5一個是負責18S rRNA m6A修飾的酶,并確認ZCCHC4為28S rRNAm6A修飾酶;METTL5必須與已知的甲基轉移酶激活劑TRMT112形成異源二聚體復合物,以在細胞中獲得代謝穩定性。文章還提供了METTL5-TRMT112的第一個原子分辨率結構。

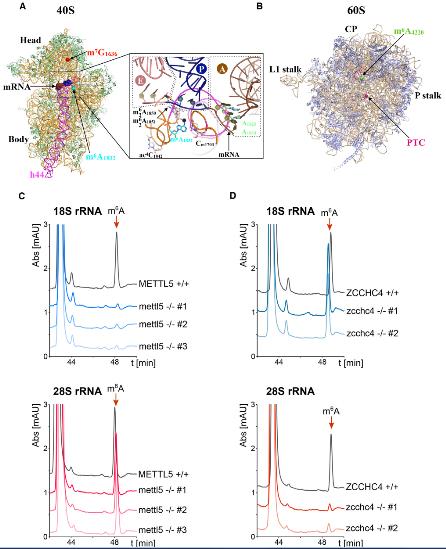
1.鑒定m6A1832和m6A4220上沉積的18S和28S rRNA甲基轉移酶
為了鑒定在18S rRNA的A 1832和28S rRNA的A 4220處沉積m 6 A 的甲基轉移酶,在UniProt數據庫中搜索了具有N6-腺苷特異性DNA甲基轉移酶。檢索到七個結果結果:HEMK1、HEMK2、TRMT11、MGAM2、METTL4、METTL5和ZCCHC4。根據人類蛋白質圖譜計劃,METTL4是人類U2-OS細胞中線粒體蛋白;METTL5和ZCCHC4分別位于核仁和細胞核,即核糖體生物發生的初始步驟。有研究報道,人類METTL5與RNA特異性結合,但不與DNA結合。這些結果促使作者研究了METTL5和ZCCHC4是否可能是導致人類rRNA上m 6 A沉積的酶。
為了驗證這一假設,通過CRISPR-Cas9基因編輯消除了人類細胞(HCT116)二倍體兩個等位基因上蛋白質的部分編碼序列,從METTL5中刪除了外顯子3,從ZCCHC4中刪除了外顯子7。三個獨立的mettl5-/-細胞系和兩個獨立的zcchc4-/-細胞系,通過PCR驗證基因組DNA上兩個等位基因的缺失。為了測試METTL5和ZCCH4在核糖體RNA修飾中的作用,通過離心從不同的mettl5-和zcchc4缺失的細胞系以及對照HCT116細胞中純化了成熟的18S和28S rRNA,然后將其消化成核苷并用HPLC定量分析。在對照細胞中,m6A核苷很容易在純化的18s和28s中檢測到,48分鐘洗脫。在所有三個mettl5-/-細胞中,18S rRNA m6A峰丟失,而28S rRNA m6A峰保持不受影響。相反,在兩個zcchc4-/-克隆中,18S rRNA m6A峰不變,而28S rRNA m6A峰消失。這些數據可以得出結論, METTL5和ZCCHC4是人類甲基轉移酶,分別負責在18S rRNA上形成m 6 A 1832和在28S rRNA 上形成m 6 A 4220。ZCCHC4鑒定為28S rRNA m6A修飾酶與最近的文獻報道完全一致。
2.METTL5和ZCCHC4可作用于細胞生長和核糖體生物發生
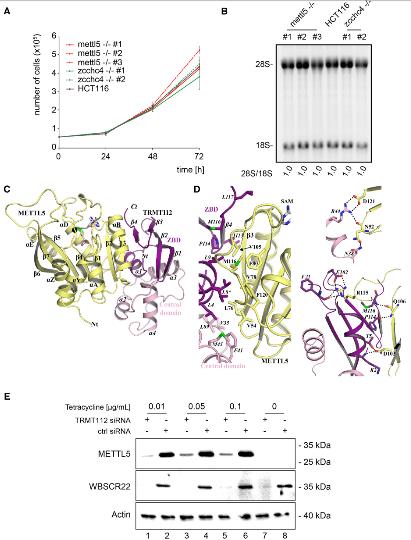
確定了18S和28S rRNA m 6 A甲基化酶后,檢查這些修飾是否對細胞生長和核糖體生物發生有影響。首先,利用活體細胞染色計數監測細胞增殖,實驗結果表明無METTL5或ZCCHC4基因的對細胞生長沒有顯著影響。接下來,從HCT116細胞和ETTL5-/-和ZCCHCC4-/-細胞中提取的總RNA進行瓊脂糖凝膠電泳和Ethylenediamine tetramine染色,以顯示成熟的18S和28S rRNA。發現缺失METTL5或ZCCHC4基因的細胞正常產生成熟的rRNA。通過定量高分辨率Northern印跡進行的詳細的pre-rRNA加工分析顯示,所有主要的pre-rRNA種類均帶有特異性探針,并且沒有重大差異。總之,METTL5是18S rRNA m6A裝飾酶,ZCCHC4 是28S rRNAm6A裝飾酶,這兩種酶都不是細胞生長或成熟rRNA產生所必需的,也不是任何沉積的修飾。
3.人METTL5與TRMT112形成異源二聚體,獲得代謝穩定性
在作者最近的研究中,旨在描述古細菌中甲基轉移酶激活劑Trm112的相互作用網絡。發現METTL5同源基因HVO _1475(以下簡稱HVO Mettl5,35%的序列同一性METTL5),這導致測試HvoMettl5是否可以直接與HvoTrm112相互作用,并由此擴展人類METTL5是否可能與TRMT112相互作用。Hvo Mettl5與Hvo Trm112發生物理相互作用,而人類METTL5與TRMT112直接相關。當ZCCHC4與TRMT112共表達時,沒有觀察到穩定作用。通過X-射線衍射確定蛋白晶體結構。人TRMT112包含兩個結構域:鋅結合域(ZBD)和中央域。
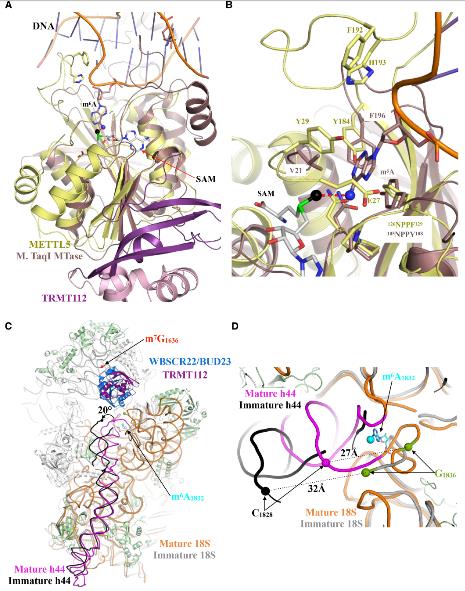
4.METTL5-TRMT112與DNA和RNAm6A甲基轉移酶對核糖體生物發生的意義比較
METTL5-TRMT112的結構是繼mElt3-METTL14,METL16和CAPAM之后的人類確定的第四個RNA m6A甲基轉移酶結構。沒有觀察到METTL5-TRMT112與上述三種甲基轉移酶有明顯的相似性。這表明METTL5-TRMT112可能以其自己的特定方式運行。有趣的是,將METTL5-TRMT112的結構與結合到短雙鏈DNA片段的m6A DNA甲基轉移酶的結構進行比較時,發現了意想不到的共性。一個DNA鏈的目標腺嘌呤環從雙螺旋中伸出并指向活性位點,并由105NPPY108標記以及Val21和Phe196側鏈協調。這些分別對應于人METTL5的126NPPF129,Tyr29和Tyr184,這可能像M. TaqI一樣在其活性位點目標為腺嘌呤,最佳地定位腺嘌呤以將甲基基團轉移到其N6原子上。
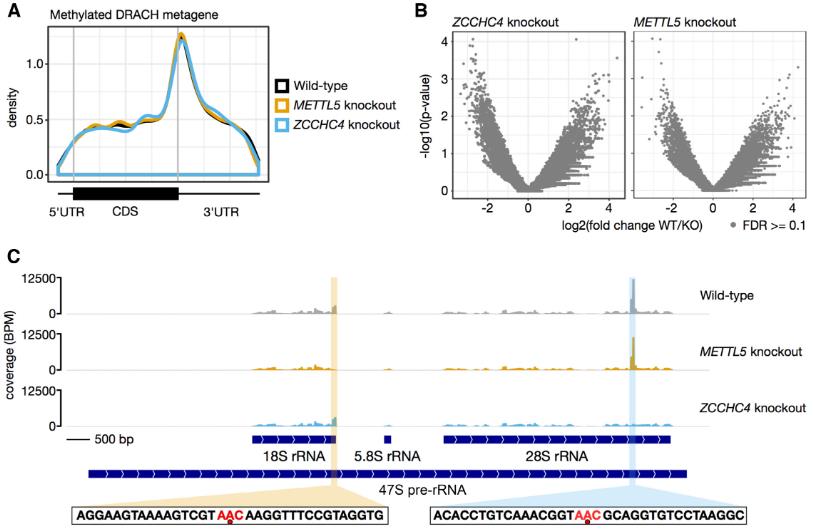
5.METTL5和ZCCHC4僅是m6A核糖體RNA的編輯器
我們想知道是否METTL5-TRMT112或ZCCHC4可以進行動作在核糖體RNA以外的RNA底物上。為了驗證這一問題,使用miCLIP技術對缺少METTL5或ZCCHC4的人類細胞的轉錄組進行m6A的單核苷酸圖譜。雖然12346個m6A位點重新定位在同基因對照細胞的mRNA上;在缺少METTL5或ZCCHC4的情況下,mRNA或lncRNA中的位點差異很大。miCLIP分析進一步證實,METTL5-TRMT112負責18S rRNA m6A 1832安裝,ZCCHC4用于28S rRNA m6A 4220沉積。總之,METTL5和ZCCHC4分別對18S和28S rRNA m6A修飾具有高度特異性。
結論:
METTL5一個是負責18S rRNA m6A修飾的酶,并確認ZCCHC4為28S rRNAm6A修飾酶;METTL5必須與甲基轉移酶激活劑TRMT112形成異源二聚體復合物,以在細胞中獲得代謝穩定性。并且確定了METTL5-TRMT112的第一個原子分辨率結構。