






你真的認(rèn)識(shí)lncRNA嗎?
導(dǎo)語(yǔ):
現(xiàn)在關(guān)于lncRNA的研究已經(jīng)不在少數(shù),關(guān)于它的功能大家也都一定的了解,從哺乳動(dòng)物基因組轉(zhuǎn)錄出多種不能編碼蛋白質(zhì)的lncRNA,它們通過控制細(xì)胞核中的核結(jié)構(gòu)和轉(zhuǎn)錄以及通過調(diào)節(jié)細(xì)胞質(zhì)中的mRNA穩(wěn)定性,翻譯和翻譯后修飾而成為基因表達(dá)網(wǎng)絡(luò)中的重要調(diào)節(jié)劑。但是細(xì)胞中l(wèi)ncRNA的功能你是否已經(jīng)完全掌握了呢,不如和我們一起看看這篇文獻(xiàn)所做的細(xì)胞中l(wèi)ncRNA的功能總結(jié)吧!
參考文獻(xiàn):
Yao Run-Wen,Wang Yang,Chen Ling-Ling,Cellular functions of long noncoding RNAs.[J] .Nat. Cell Biol., 2019, 21: 542-551. IF:17.728
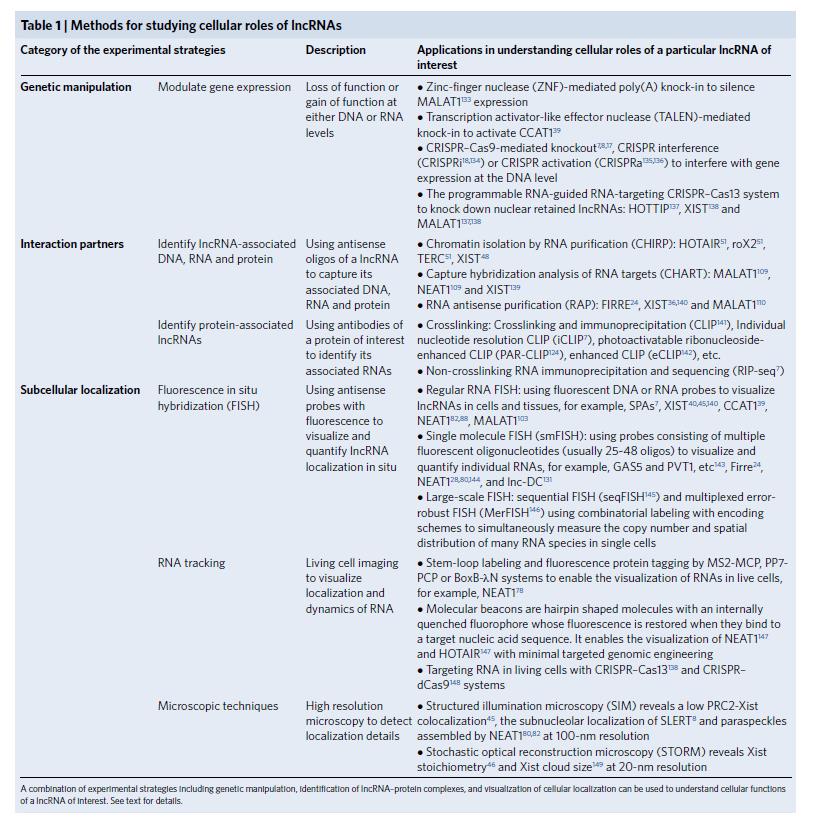
圖1:細(xì)胞中l(wèi)ncRNA的功能研究的常用方法
細(xì)胞中l(wèi)ncRNA的功能主要分為6種:1、lncRNA在細(xì)胞核內(nèi)的功能2、lncRNA在染色質(zhì)結(jié)構(gòu)中的作用3、 lncRNA在染色質(zhì)重建中的作用4、 lncRNA在轉(zhuǎn)錄調(diào)控中的作用5、 lncRNA在核體調(diào)節(jié)中的作用6、lncRNA在細(xì)胞質(zhì)中的功能。
1、 lncRNA在細(xì)胞核內(nèi)的功能
lncRNA多數(shù)定位在細(xì)胞核, 是因?yàn)閘ncRNA上的Alu元件表現(xiàn)出C富集,是靈長(zhǎng)類特異性的短散布核元件(SINE),通過與核基質(zhì)蛋白HNRNPK的結(jié)合促進(jìn)lncRNA核保留。lncRNA上含有的與核蛋白相關(guān)的順式元件可以防止核蛋白流出細(xì)胞核,并成為核組織和功能的調(diào)節(jié)者。
2、 lncRNA在染色質(zhì)結(jié)構(gòu)中的作用
為協(xié)調(diào)轉(zhuǎn)錄調(diào)控,間期染色體結(jié)構(gòu)嚴(yán)密。如圖2所示,紅色的lncRNA可以在不同水平調(diào)節(jié)基因組組織。
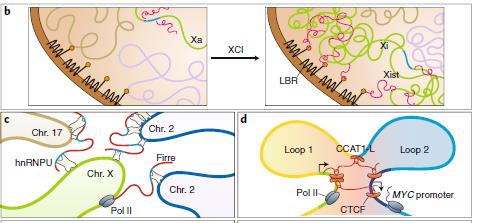
圖2 b:lncRNA調(diào)節(jié)染色體的結(jié)構(gòu)c,d: lncRNA調(diào)節(jié)不同染色體之間和染色體內(nèi)部的相互作用。XCI:X染色體失活,Xi:無(wú)活性X染色體,Xist:X染色體特異性失活轉(zhuǎn)錄因子,LBR:核纖層蛋白受體,F(xiàn)irre:從X染色體轉(zhuǎn)錄并逃脫X染色體失活的RNA,可與hnRNPU結(jié)合
3、lncRNA在染色質(zhì)重建中的作用
許多核定位的lncRNA,無(wú)論是順式或反式,都與染色質(zhì)及染色質(zhì)重建有關(guān)。Xist與維管介質(zhì)直接相互作用,用于為HCOOH和甲狀腺激素受體(SMART)/組蛋白去乙酰化酶1(HDAC1)相關(guān)的阻遏蛋白。 SHARP反過來(lái)招募SMART及其相互作用的HDAC3,導(dǎo)致組蛋白去乙酰化和轉(zhuǎn)錄抑制。染色質(zhì)調(diào)節(jié)蛋白質(zhì)逐步募集到未來(lái)的Xi最終導(dǎo)致廣泛的轉(zhuǎn)錄沉默。
Mhrt(肌球蛋白重鏈相關(guān)RNA轉(zhuǎn)錄物)是源自肌球蛋白重鏈7基因的反義轉(zhuǎn)錄的可變剪接的細(xì)胞核保留的lncRNA簇。 Mhrt通過拮抗Brg1保護(hù)心臟免于病理性肥大,Brg1是已知促進(jìn)病理性心臟肥大的BAF染色質(zhì)重塑復(fù)合物的催化亞基。Mhrt通過與RNA解旋酶結(jié)構(gòu)域(Brg1和 DNA結(jié)合位點(diǎn))相關(guān)聯(lián)將Brg1從靶基因組基因座上隔絕。
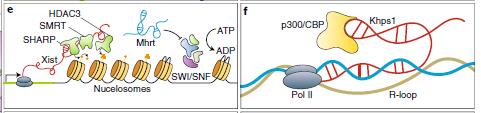
圖3:lncRNA促進(jìn)或抑制染色質(zhì)修飾體的聚集SMART:維HCOOH和甲狀腺激素受體的沉默介質(zhì),HDAC1 :組蛋白去乙酰化酶1,SHARP: HDAC1相關(guān)的阻遏蛋白,Mhrt:肌球蛋白重鏈相關(guān)RNA轉(zhuǎn)錄
4、 lncRNA在轉(zhuǎn)錄調(diào)控中的作用
lncRNA可以通過形成R-環(huán)結(jié)構(gòu)聚集轉(zhuǎn)錄因子或通過干擾靶向基因座處的Pol II轉(zhuǎn)錄直接調(diào)節(jié)轉(zhuǎn)錄。一些反義lncRNA通過形成R-環(huán)(具有與雙鏈DNA雜交的RNA的三鏈核酸結(jié)構(gòu))調(diào)節(jié)正義mRNA轉(zhuǎn)錄。 R環(huán)的局部形成可以使順式連接lncRNA并將轉(zhuǎn)錄輔因子募集到相應(yīng)的啟動(dòng)子區(qū)域。例如,Khps1以反義方向轉(zhuǎn)錄為原癌基因SPHK1。將Khps1連接到SPHK1轉(zhuǎn)錄起始位點(diǎn)(TSS)上游的同型嘌呤拉伸導(dǎo)致形成R-環(huán),其將組蛋白乙酰轉(zhuǎn)移酶p300 / CBP錨定在SPHK1啟動(dòng)子上(圖4f)。
在啟動(dòng)子Pol II復(fù)合物中參入Alu RNA并在純化的Pol II轉(zhuǎn)錄系統(tǒng)和細(xì)胞中阻斷轉(zhuǎn)錄起始(圖4g)。 Alu RNA含有兩個(gè)松散的結(jié)構(gòu)域,每個(gè)結(jié)構(gòu)域結(jié)合一個(gè)Pol II分子對(duì)轉(zhuǎn)錄抑制至關(guān)重要。一旦從預(yù)啟動(dòng)復(fù)合物上移除Alu,轉(zhuǎn)錄恢復(fù)。
轉(zhuǎn)錄延伸也由lncRNA調(diào)節(jié)。與Alu和B2 RNA一樣,7SK RNA是Pol III轉(zhuǎn)錄的另一種豐富的RNA。它與陽(yáng)性轉(zhuǎn)錄延伸因子b(P-TEFb)結(jié)合并抑制其激酶活性,這是Pol II延伸65所需的(圖4g)。作為7SK RNA-蛋白質(zhì)復(fù)合物的一部分,DEAD-box RNA解旋酶DDX21以RNA解旋酶依賴性方式促進(jìn)P-TEFb從該抑制復(fù)合物中的釋放,導(dǎo)致C-末端結(jié)構(gòu)域的Ser2的磷酸化增加(圖4g)。
RNA聚合酶I(Pol I)轉(zhuǎn)錄也可以由lncRNA調(diào)節(jié)。例如,SLERT根據(jù)其snoRNA末端從其轉(zhuǎn)錄位點(diǎn)轉(zhuǎn)移到細(xì)胞核中。在細(xì)胞核中,SLERT與DDX21相互作用,DDX21以不依賴于RNA解旋酶的方式結(jié)合并抑制Pol I.。SLERT結(jié)合改變DDX21構(gòu)象并釋放DDX21和Pol I機(jī)制之間的抑制性相互作用,從而促進(jìn)rDNA轉(zhuǎn)錄(圖4h)。
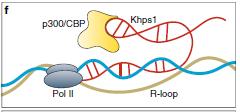
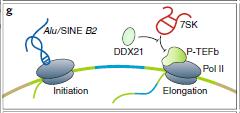
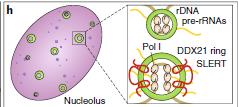
圖4:lncRNA可以通過形成R-環(huán)結(jié)構(gòu)聚集轉(zhuǎn)錄因子或通過干擾靶向基因座處的Pol II轉(zhuǎn)錄直接調(diào)節(jié)轉(zhuǎn)錄 g:lncRNA干擾Pol II聚合體。P-TEFb:陽(yáng)性轉(zhuǎn)錄延伸因子b,DDX21 : DEAD-box RNA解旋酶f:lncRNA通過形成R環(huán)調(diào)節(jié)轉(zhuǎn)錄。Khps1:原癌基因SPHK1反義轉(zhuǎn)錄的lncRNA,p300 / CBP :組蛋白乙酰轉(zhuǎn)移酶h:轉(zhuǎn)錄或lncRNA基因座調(diào)節(jié)轉(zhuǎn)錄。SLERT:snoRNA末端的lncRNA增強(qiáng)前體核糖體RNA轉(zhuǎn)錄
5、 lncRNA在核體調(diào)節(jié)中的作用
核體(NBs)是動(dòng)態(tài)的無(wú)膜RNA-蛋白質(zhì)復(fù)合物。 lncRNA調(diào)節(jié)NB的完整性和功能,在轉(zhuǎn)錄后水平改變的基因表達(dá)。
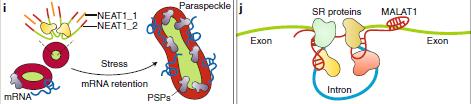
圖5:lncRNA在核體調(diào)節(jié)中的作用Paraspeckle:“核斑”,NEAT1:核內(nèi)富集分轉(zhuǎn)錄本1,MALAT1:肺癌轉(zhuǎn)移相關(guān)轉(zhuǎn)錄本1
6、lncRNA在細(xì)胞質(zhì)中的功能
許多l(xiāng)ncRNA被輸出到細(xì)胞質(zhì),在細(xì)胞質(zhì)中調(diào)節(jié)mRNA穩(wěn)定性,調(diào)節(jié)翻譯并干擾翻譯后修飾(PTM)。
lncRNAs可以通過多種方式影響mRNA的轉(zhuǎn)換。首先,它們可以通過相關(guān)的miRNA調(diào)節(jié)mRNA穩(wěn)定性。Cyrano也與miR-7結(jié)合,但這種lncRNA不是隔離miR-7,而是通過促進(jìn)3'末端的拖尾和修剪來(lái)促進(jìn)其有效破壞,從而誘導(dǎo)靶向RNA指導(dǎo)的miRNA降解,導(dǎo)致Cdr1在大腦中積累。 Cyrano缺乏使miR-7積累,導(dǎo)致Cdr1在神經(jīng)元中降解(圖6k)。這些研究結(jié)果表明,ceRNA理論比以前認(rèn)為的要復(fù)雜得多,不同類型的ncRNA可以協(xié)同工作以建立復(fù)雜的監(jiān)管網(wǎng)絡(luò)。其次,lncRNAs可通過募集蛋白質(zhì)來(lái)降解mRNA來(lái)調(diào)節(jié)mRNA的穩(wěn)定性。 例如,一組含有Alu的lncRNA可以反式激活Staufen 1(STAU1)介導(dǎo)的mRNA衰變(SMD)。 靶向SMD的mRNA在3'-UTR內(nèi)含有Alu元件,其可以與lncRNA中的互補(bǔ)Alu堿基配對(duì)以形成雙鏈RNA(dsRNA),這是STAU1識(shí)別的結(jié)構(gòu)。(圖61)。第三,lncRNA可以作為參與mRNA衰變的RBP的分子誘餌起作用。 NORAD是哺乳動(dòng)物中豐富且保守的lncRNA,在細(xì)胞質(zhì)中充當(dāng)PUMILIO 1和PUMILIO 2(PUM1 / 2)的儲(chǔ)庫(kù),以限制它們靶向mRNA降解的可用性(圖61)。
lncRNA參與翻譯調(diào)控。例如,lincRNA-p21與HuR相互作用,這種結(jié)合有利于let-7 / Ago2的募集使lincRNA-p21不穩(wěn)定。在喪失HuR后,lincRNA-p21通過堿基配對(duì)積累并與JUNB和CTNNB1 mRNA結(jié)合,通過募集翻譯阻遏物Rck129來(lái)抑制它們的翻譯(圖6m)。lncRNAs也可以激活mRNA翻譯。幾種lncRNA通過掩蔽由PTM酶或PTM位點(diǎn)結(jié)合的位點(diǎn)來(lái)調(diào)節(jié)PTM。 lnc-DC調(diào)節(jié)STAT3的磷酸化,STAT3是控制DC分化的TF131。Lnc-DC通過阻止蛋白酪氨酸磷酸酶SHP1的結(jié)合直接結(jié)合STAT3并促進(jìn)Tyr705的磷酸化(圖6n)。
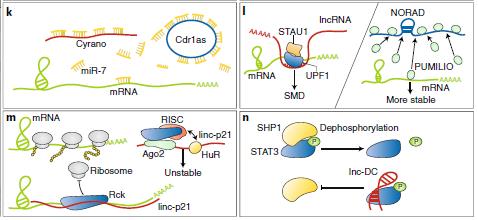
圖6:lncRNA在細(xì)胞質(zhì)中的功能Cdr1as:小腦變性相關(guān)蛋白1反義轉(zhuǎn)錄的circRNA,Cyrano :一種 lncRNA,STAU1 : Staufen 1 蛋白,SMD : STAU1 介導(dǎo)的mRNA降解,NORAD :由DNA損傷激活的lncRNA,RISC:RNA誘導(dǎo)沉默復(fù)合體,Lnc-DC:在人常規(guī)樹突細(xì)胞質(zhì)中專一表達(dá)lncRNA