






ScRNA-seq+免疫:前列腺癌淋巴結轉移相關的免疫抑制
轉移是導致前列腺癌(PCa)患者死亡的疾病進展的關鍵。本文旨在通過使用單細胞RNA測序(scRNA-seq)分析PCa中腫瘤微環境(TME)的異質性來探討淋巴結轉移(LNM)的機制。從四個PCa組織樣本中獲得32,766個細胞進行scRNA-seq,結果顯示,在LNM中只有EEF2+和FOLH1+腔亞群存在,并且它們出現在腔細胞分化的初期階段,這通過驗證實驗證實。MYC通路在EEF2+和FOLH1+腔亞群中富集,MYC與PCa LNM相關。此外,MYC不僅促進PCa的進展,還通過調節PDL1和CD47導致TME的免疫抑制。在LNM中,TME中CD8+T細胞的比例以及NK細胞和單核細胞中的比例較原發病灶中較低,而Th和Treg細胞相反。此外,TME中的這些免疫細胞經歷了轉錄重編程,包括CCR7+和IL7R+的CD8+T亞群,以及表達腫瘤相關基因的M2樣單核細胞亞群,如CCR7、SGKI和RPL31。此外,與腫瘤進展、腫瘤代謝和免疫抑制密切相關的STEAP4+、ADGRF5+和CXCR4+、SRGNC+纖維母細胞亞群表明它們在PCa轉移中的貢獻。同時,通過多色免疫熒光證實了PCa中存在CXCR4+纖維母細胞。總之,PCa LNM中腔細胞、免疫和間質細胞的顯著異質性可能不僅直接促使腫瘤進展,還間接導致TME免疫抑制,這可能是PCa轉移的原因,其中MYC發揮了作用。本文于2023年5月發表在《Experimental Hematology & Oncology》IF:10.9期刊上。
技術路線

主要實驗結果:
1、原發病灶和淋巴結轉移中的TME細胞組成
對兩名患有PCa的患者的四個腫瘤樣本進行scRNA-seq分析,以確定它們的細胞組成(圖1A)。共獲得32,766個細胞的單細胞轉錄組,其中16,717個細胞來自原發病灶,16,049個細胞來自淋巴結轉移。UMAP分析基于細胞的基因型和典型標記基因,鑒定了14個主要細胞類群(圖1B和D),包括:(1)具有GZMK、CD8A和IFNG高表達的CD8+T細胞;(2)特異性表達KRT18和EPCAM的腔細胞;(3)具有ACTA2和TAGLN高表達的纖維母細胞;(4)具有GZMB、NKG7和GNLY高表達的NK細胞;以及(5)特異性表達HLA-DRA、C1QA和C1QB的單核細胞,以及其他細胞類型的標記基因(圖1D)。結果顯示了每個細胞類型中前10個高表達基因的表達譜,以及每個細胞類型的細胞比例(圖1C和E)。值得注意的是,每個原發病灶樣本中都存在所有細胞類型,而在轉移病灶中基底細胞和中性粒細胞幾乎不存在(圖1C)。TME細胞組成的顯著差異暗示著原發病灶和淋巴轉移之間明顯的異質性。
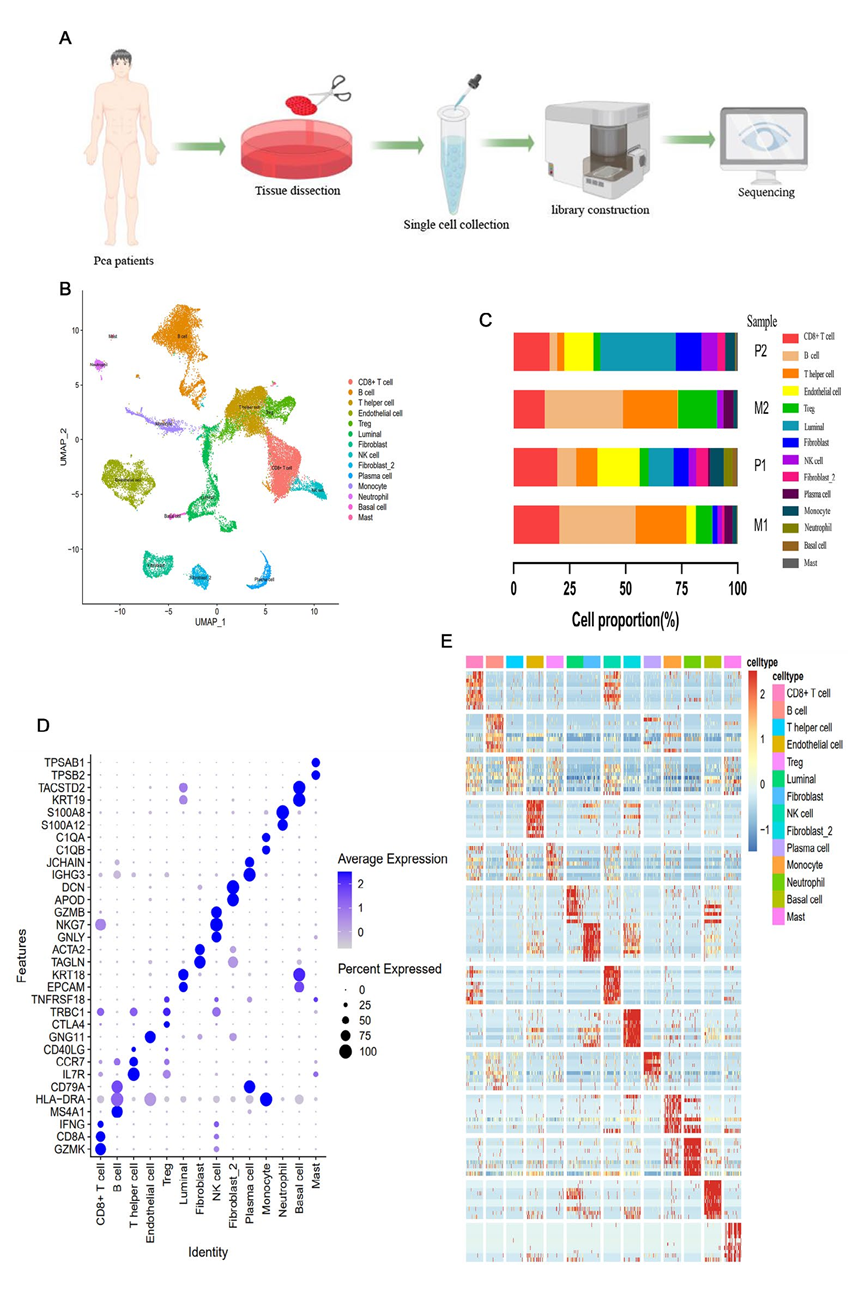
圖1 通過scRNA-seq鑒定出PCa中的14種細胞類型
2、腔細胞的轉錄異質性和InferCNV
根據先前的研究,腔細胞和基底細胞是前列腺癌可能的起始細胞。對腔細胞的UMAP分析確定了總共七個亞群(圖2A),并展示了不同腔細胞亞群的基因表達模式(圖2B)。還展示了每個腔細胞亞群中前五個標記基因,分別是EEF1A2、HBB、IGKC、NPY和FOLH1在腔上1中高表達。在腔上5中HBB和IGKC高表達,而NPY和FOLH1在腔上7中高表達(圖2C)。GSVA顯示MYC和氧化磷酸化信號通路在腔上1亞群中富集,而TNF-α信號通路活性在腔上3亞群中增強。此外,腔上4亞群具有高的蛋白分泌和雄激素應答信號通路富集得分,而E2F和G2M信號通路在腔上6亞群中富集。腔上7亞群展示了在血管生成信號通路中的增強活性(圖2D)。
InferCNV算法被用于識別染色體CNAs,以進一步驗證七個腔上亞群中是否存在惡性細胞。根據先前的研究,該方法可以識別典型的PCa基因改變,包括染色體8q增益和染色體8p、13和16q缺失。此外,先前的DNA測序研究揭示0-50%的PCa基因組具有CNAs,并且CNAs也是PCa的預后因素。而且,在大多數PCa病例中,次克隆CNAs的負擔遠遠超過克隆負擔。本文的InferCNV結果顯示在圖2E中。腔上亞群1和5以及3和7中的所有細胞都被聚集在一起。其他亞群中的所有細胞都被單獨分組(圖2E)。在CNAs的熱圖中,紅色和藍色分別表示片段染色體中基因表達值過高和過低。這意味著紅色顏色越深,染色體擴增的程度越高,而藍色顏色越深,染色體喪失的程度越高。因此,研究結果顯示腔上亞群1和5以及3和7中的染色體CNAs的程度遠高于其他簇。因此,腔上亞群1和5以及3和7中的細胞可能是惡性細胞(圖2E)。此外,腔上亞群1和5僅出現在轉移病灶中,暗示這兩個亞群可能在惡性細胞中包含有轉移能力的細胞(圖2G)。
進一步分析了原發病灶和淋巴結轉移之間的差異表達基因(DEGs)及其功能富集,以揭示PCa淋巴結轉移中的特定基因表達模式(圖2H-I)。結果顯示主要富集的功能性DEG途徑與免疫相關,如白細胞和淋巴細胞活性的調控以及B細胞活性(圖2I),表明腫瘤免疫可能對PCa的淋巴結轉移產生影響。此外,還分析了腔細胞的轉錄調控因子在原發病灶和淋巴結轉移之間的差異(圖2F和J)。JDP2、IRF1、ENO1、HOXB2、IRF8、NELFE、ESRRA、SP8、KLF9、SF1、TFAP2A、IRF7、CREB3、GTF2F1、MXI1、CREB5、EGR3、FOXO3、NFATC3和CREM在淋巴結轉移中都被激活,而在原發病灶中沒有激活(圖2F和J)。特別是NELFE在淋巴結轉移中的表達顯著高于原發病灶。有報道稱NELFE在許多癌癥中能促進侵襲和轉移。因此,上述結果表明這些轉錄因子可能參與了PCa的淋巴結轉移(圖2J)。
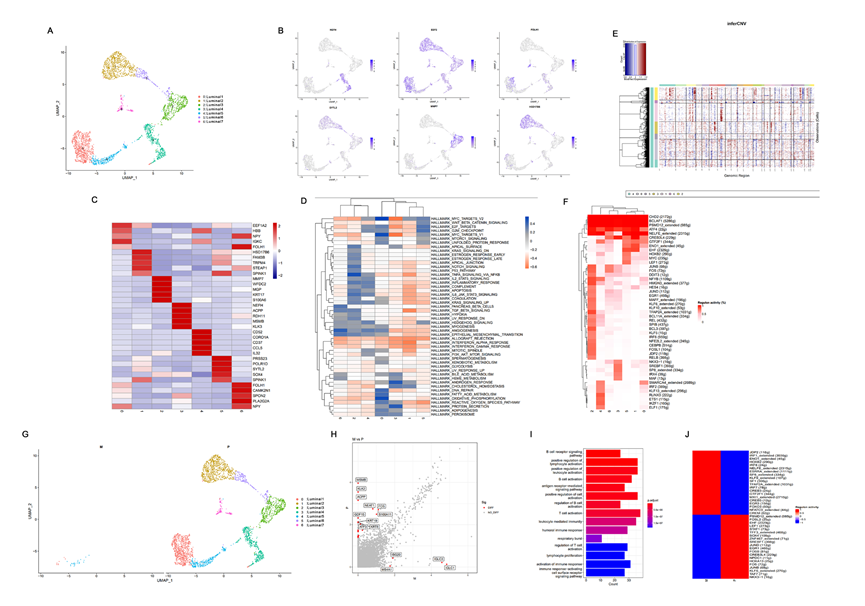
圖2 腔室細胞的異質性和CNV分析
隨后,選擇在EEF2+和FOLH1+腔上亞群中具有特異性的標記基因EEF1A2和CCL5進行免疫組化染色在PCa組織芯片上,結果在正常組織中顯示陰性結果。在原發病灶和淋巴結轉移中的一些前列腺癌細胞呈現陽性表達,這表明在PCa中存在這兩個亞群的細胞(圖3A)。免疫熒光顯示MEEF1A2和CCL5在LNCap細胞中均有表達(圖3B)。在LNCap細胞中下調MEEF1A2和CCL5的表達可以顯著降低細胞增殖和轉移的能力(圖3C-F)。
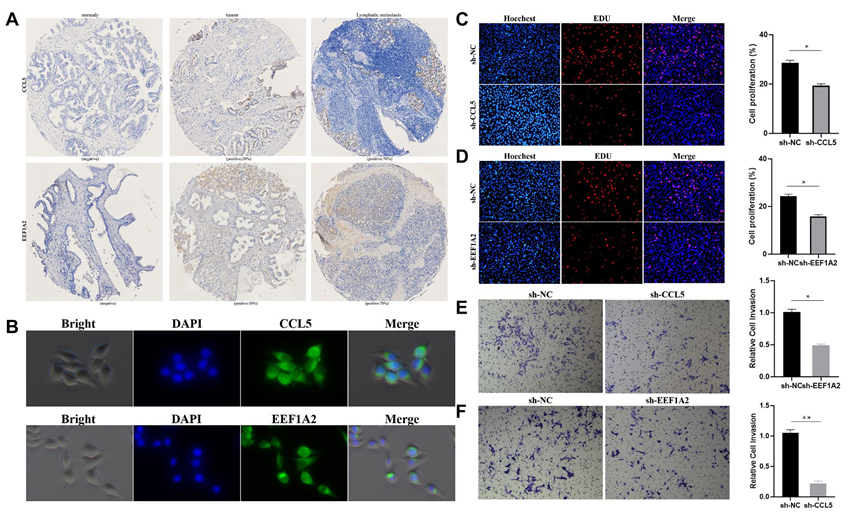 圖3 PCa中存在EEF2+和FOLH1+腔上亞群細胞。
圖3 PCa中存在EEF2+和FOLH1+腔上亞群細胞。
3、MYC可能通過促進腫瘤細胞進展和免疫抑制來促使淋巴結轉移
腔上亞群1和5出現在淋巴結轉移中。MYC通路在這些細胞中也顯示富集,因此進行進一步的調查。芯片IHC結果顯示MYC在細胞核和細胞質中表達,并且在PCa組織中的表達顯著強于正常前列腺組織。重要的是,淋巴結轉移中的MYC表達顯著高于原發病灶(圖4A),表明MYC與PCa轉移有關。免疫熒光結果顯示,LNCap細胞中MYC、PDL1和CD47均有表達,而PDL1和CD47主要表達在細胞質中(圖4B)。從NCBI和JASPAR數據庫獲取的結果顯示,PDL1和CD47的啟動子區域有MYC的結合位點,證明PDL1和CD47是MYC的靶基因(圖4C)。通過敲低LNCap細胞中MYC的表達后,發現PDL1和CD47的mRNA和蛋白水平顯著下調(圖4D-F)。此外,Transwell結果表明MYC表達下調顯著降低了LNCap細胞的轉移能力(圖4G-H)。
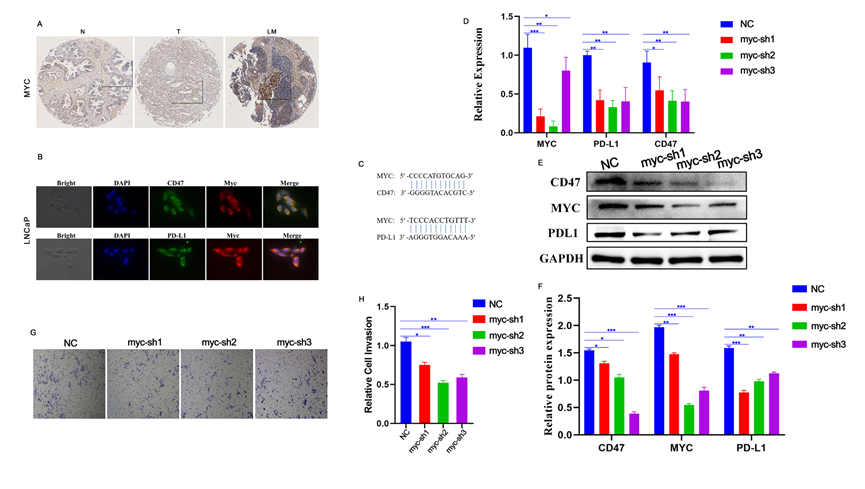
圖4 MYC通過CD47和PD-L1驅動腫瘤進展
4、腔細胞軌跡分析
細胞軌跡結果顯示在圖4中,其中Monocle 2將所有腔細胞分為七個狀態(圖5B)。圖4A顯示了根據Seurat進行的降維聚類后,細胞根據其簇的分化軌跡。簇0和4分布在圖的右側,而簇2和6分布在圖的左側。另外的三個簇(1、3和5)位于圖中部的下方(圖5A)。此外,腔細胞的分化軌跡以第二分支點為重要的轉折點,從圖的右側到左側和下部(圖5C)。因此,腔細胞的總體分化發生在簇0、1和4到簇2、3和6之間。InferCNV結果顯示簇0、2、4和6是潛在的惡性細胞,而簇1、3和5是非惡性細胞。兩個簇0和4是轉移細胞,暗示具有轉移能力的癌細胞早期發生(圖5C)。此外,惡性細胞的分化方向與非惡性細胞不同。
接下來,探索了腔細胞分化過程中的轉錄組變化。選擇具有不同表達特征的代表性基因,觀察它們在細胞分化過程中的動態變化(圖5C)。MYC相關的鋅指蛋白(MAZ)在一些重要基因,包括MYC、RAS和CT-1中發揮轉錄調控作用。MAZ的表達在細胞分化時間內首先增加,然后急劇下降(圖5D)。唾液酸蛋白(SPN),也稱為LSN和CD43,是一種跨膜唾液酸糖蛋白。隨著腔細胞的分化,SPN基因表達在基線處最初保持不變,然后急劇增加,稍有下降,最終迅速增加(圖5D)。圖5E顯示了在腔細胞分化過程中特征基因表達變化的可視化和聚類結果。隨后,根據分化過程,將腔細胞分為命運1、分支前和命運2子集。在腔細胞從命運1到命運2的分化過程中,所有變化的基因可以根據它們的表達水平分為四類(圖5F)。
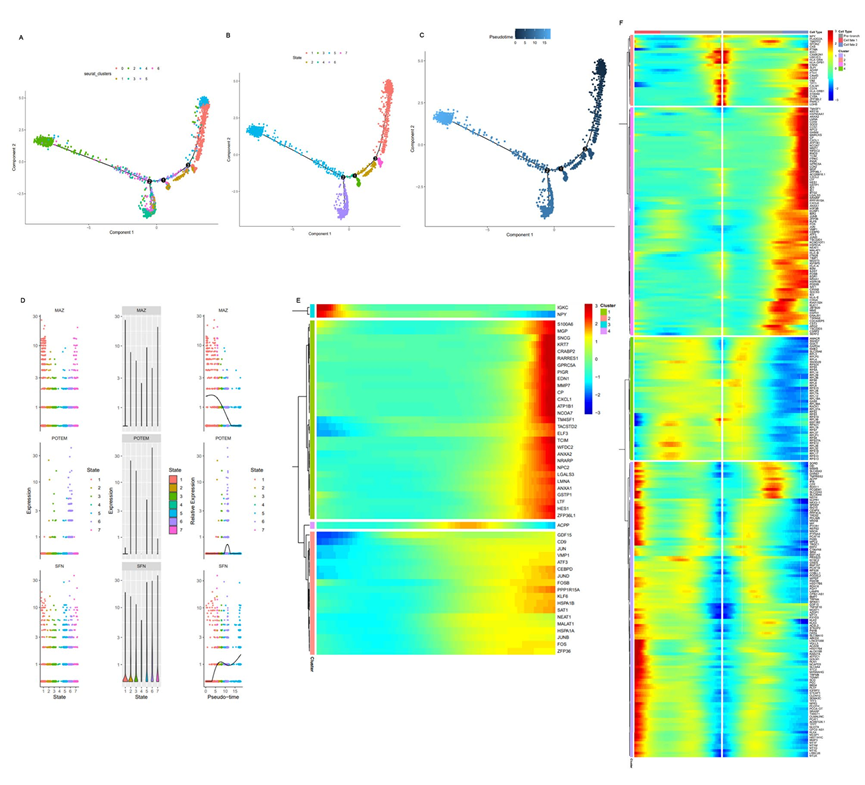
圖5腔內細胞的軌跡分析
5、原發病灶和淋巴結轉移病灶中CD8+ T細胞的異質性特征
在本研究中,鑒定了5,843個CD8+ T細胞(原發病灶中的3,069個,轉移病灶中的2,774個)。通過CD8+ T細胞的UMAP分析鑒定了四個亞群(圖6A)。此外,結果顯示在轉移病灶中,亞群0和3的比例顯著高于原發病灶(圖6B)。而且,在標記基因中,CCR7僅在亞群0和3中表達(圖6C)。研究發現CCR7通過促進腫瘤細胞增殖和轉移、鼓勵蛋白酶分泌以及誘導血管生成和免疫抑制來促進腫瘤發展,與本文發現一致,即CD8+ T和其他腫瘤免疫細胞水平降低并經歷轉錄重組,導致淋巴結轉移中出現明顯的免疫抑制。此外,GSVA結果顯示,亞群0和3富集的通路包括Hedgehog信號、Notch信號、血管生成和MYC通路,與腫瘤增殖和發展相關(圖6F)。這些結果表明,亞群0和3表達的標記基因具有腫瘤基因的特征,可以促進腫瘤增殖和轉移,同時引起腫瘤免疫抑制。亞群2的比例在轉移病灶中也顯著小于原發病灶(圖6B),NR4A3、DUSP4和RGS1是表達較高的標記基因(圖6C)。亞群2富集的通路包括凋亡、上皮間質轉化(EMT)、氧化磷酸化和DNA修復(圖6F)。這些結果證實了亞群2中表達的標記基因,如NR4A3,可以通過介導MT-2信號通路抑制前列腺癌轉移,顯示了與CD8+ T細胞亞群0和3相反的功能。這些結果表明,TME中T細胞的異質性和免疫抑制可能是前列腺癌淋巴結轉移的重要原因。對比之前的原發病灶和淋巴結轉移病灶之間的DEGs的功能富集通路分析顯示,這些基因主要集中在細胞質翻譯、T細胞激活、正調控白細胞激活、免疫應答調節、白細胞黏附等與免疫相關的通路中(圖6D和E)。這些結果進一步證明了前列腺癌淋巴結轉移中CD8+ T細胞具有促使腫瘤進展并導致免疫抑制的特征。
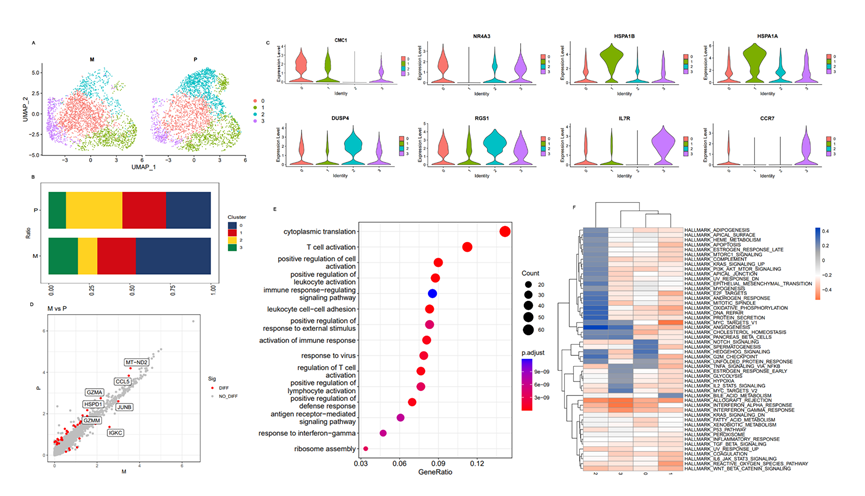
圖6 原發性和轉移性病變間CD8+T細胞的異質性分析
6、原發病灶和淋巴結轉移樣本中單核細胞的異質性
髓樣細胞是除淋巴樣細胞外TME中的另一個重要組成部分。在本研究中,共鑒定了1,203個單核細胞(原發病灶中的930個,轉移病灶中的273個),而在淋巴結轉移病灶中的單核細胞的數量和比例顯著小于原發病灶。單核細胞的UMAP分析確定了共計六個亞群(圖7A)。此外,在轉移病灶中,亞群1、2、4和5的細胞比例高于原發病灶(圖7B)。所有四個亞群中均發現BIRC3的過表達,而亞群1和2同時過表達M1特征基因CCL3(圖7C)。此外,GSVA分析顯示亞群1和2富集了TNF-α信號、炎癥反應和TGF-β信號通路。除了BIRC3之外,促進腫瘤增殖和發展的特征基因,包括RPL31和EREG,均在亞群1和2中表達(圖7C,F)。以上結果表明,亞群1和2可能是從M1到M2的過渡階段,這與Azizi等人的研究結果一致,他們發現乳腺癌中一些TAM細胞群體高度表達了M1(如CCL3)和M2(如MARCO和NRP2)的特征基因。這意味著TME中的TAM分化也是一個連續和漸進的過程,而不是傳統上認為的兩個離散的狀態。此外,GSVA顯示亞群4和5富集了E2F、MYC、G2M、TNF-α信號和Hedgehog信號通路。以上結果表明,亞群4和5具有M2的特征。然而,在原發病灶中,亞群0和3的細胞比例顯著高于淋巴結轉移,暗示其可能具有抑制腫瘤的功能。GSVA結果表明蛋白分泌、氧化磷酸化、膽汁酸代謝等與代謝相關的通路主要富集在亞群0和3中(圖7C,F),表明這些細胞可能通過調節免疫和代謝來干擾TME。此外,原發病灶和淋巴結轉移病灶之間的單核細胞DEGs進一步調查。DEGs的功能富集通路分析顯示,它們主要集中在免疫應答激活、白細胞遷移、正調控細胞激活等與免疫相關的通路中(圖7D、E)。

圖7 原發性和淋巴轉移性前列腺癌TME中骨髓細胞的異質性分析
7、纖維細胞的異質性分析
除免疫細胞外,TME還包含癌相關的纖維細胞(CAFs)、血管內皮細胞、胞外基質等非免疫細胞組分,這些也影響腫瘤免疫微環境的功能狀態。在本研究中,共鑒定了2,456個纖維細胞(原發病灶中的2,066個,轉移病灶中的390個)(表2和3)。纖維細胞的UMAP分析揭示共計五個亞群(圖8A)。在淋巴結轉移病灶中,亞群3和4的細胞比例顯著高于原發病灶,表明亞群3和4的細胞與腫瘤的進展和轉移有關(圖8B)。進一步研究這兩個亞群中表達的特征基因揭示其他亞群中共同表達的基因,如DCN、ATF3和FLNA(圖8C)。更重要的是,在亞群3和4中特異表達的STEAP4和ADGRF5顯示了CXCR4和SRGN的顯著過表達(圖8C),這與TME中的腫瘤轉移或免疫抑制密切相關。對13例帶有淋巴轉移的前列腺癌的石蠟切片進行的多色免疫熒光染色表明,CXCR4在原發腫瘤組織和淋巴結轉移病灶中均為陽性,而在轉移病灶中的陽性率高于原發病灶,這與單細胞測序的結果一致。這表明在前列腺癌TME中存在CXCR4?+?纖維細胞(圖8D)。GSVA結果顯示,亞群3和4的纖維細胞中Hedgehog信號通路、血管生成和G2M通路顯著富集(圖8G),而這些通路的激活可能導致腫瘤的進展。以上結果表明,表達STEAP4、ADGRF5、CXCR4和SRGN的纖維細胞亞群3和4經歷了由轉錄重編程引起的顯著功能改變,與腫瘤的進展和轉移、腫瘤代謝和免疫抑制密切相關,暗示著這些纖維細胞具有促進前列腺癌轉移和進展的能力。進一步分析前列腺癌原發病灶和淋巴結轉移病灶之間的DEGs及其功能富集(圖8E-F),特別是在淋巴轉移灶中高度表達的DEGs,顯示在轉移灶中高度表達的DEGs包括與免疫相關的基因(CCL21、CCL19、CCL2和MYC)、與代謝相關的基因(STEAP4、FABP4、DPT和APOE)以及與腫瘤增殖和進展相關的基因(RGS、CCN和MYC)。DEGs的功能富集分析表明,這些DEGs主要富集在細胞質翻譯、正調節細胞黏附、細胞外基質組織以及參與調控細胞外基質和免疫應答的淋巴細胞激活的正調節等過程中。總的來說,這些結果表明在PCa的TME中存在明顯的成纖維細胞異質性。一些成纖維細胞亞群表達與腫瘤和免疫相關的基因,這可能影響腫瘤的進展并重塑TME中的腫瘤細胞和免疫細胞相互作用,從而促進PCa的轉移。
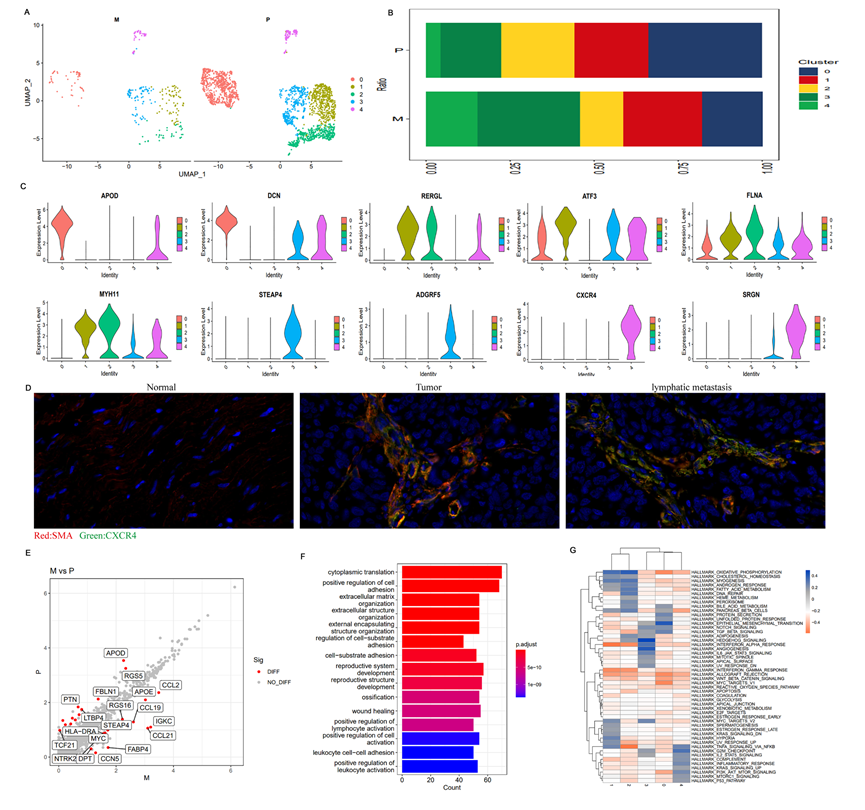
圖8 原發性和淋巴轉移性前列腺癌TME中成纖維細胞的異質性分析
8、PCa原發病變和淋巴轉移灶的細胞間通信
為確定PCa原發病變和轉移灶之間細胞間通信的差異,進行了CellChat分析。結果顯示,淋巴轉移灶中推斷的相互作用數目大于原發病變,而相互作用強度則相反。此外,在淋巴轉移灶中,間質細胞與免疫細胞之間的相互作用更為緊密,而與原發病變相比,腔細胞之間的通信減弱(圖9A、B)。對PCa中的82個激活信號通路進行了分析和鑒定,其中63個在原發病變和淋巴轉移灶中共享(圖9C、D),有9個是原發病變特有的,包括GDF、PARs、CDH、CADH、BMP、TGFb、CX3C、DESMOSOME和WNT,其中GDF和PARs更為活躍。在淋巴轉移灶中只存在的10個信號通路,包括CD22、CD45、IL16、SPP1、LIGHT、ANGPTL、GRN、ncWNT、PERIOSTIN和NEGR,除了經典的ncWNT通路可以促進腫瘤進展外,CD45和CD22最為活躍(圖9C、D)。總體而言,這些結果表明淋巴轉移灶中的細胞間通信可能導致TME中的免疫抑制狀態和腫瘤進展。
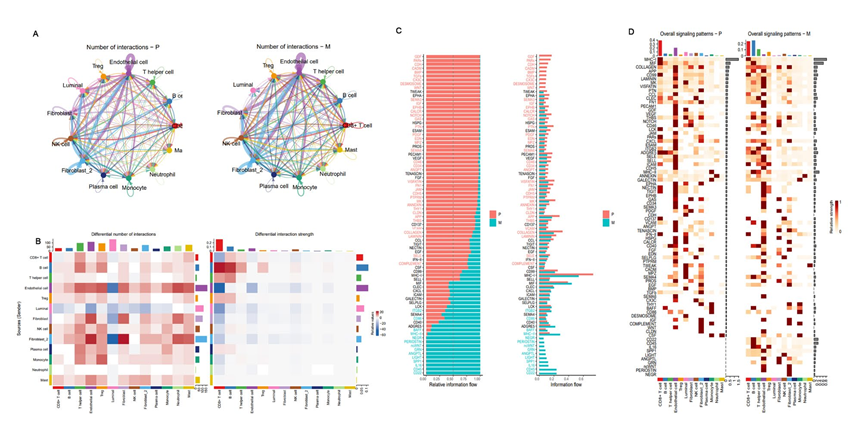
圖9原發性和淋巴轉移性前列腺癌TME的細胞間通訊
實驗方法:
組織樣本收集,細胞系培養,scRNA-seq分析,免疫組化(IHC),免疫熒光(IF),qRT-PCR,WB,Transwell實驗,EDU
參考文獻:
Xin S, Liu X, Li Z, Sun X, Wang R, Zhang Z, Feng X, Jin L, Li W, Tang C, Mei W, Cao Q, Wang H, Zhang J, Feng L, Ye L. ScRNA-seq revealed an immunosuppression state and tumor microenvironment heterogeneity related to lymph node metastasis in prostate cancer. Exp Hematol Oncol. 2023 May 23;12(1):49. doi: 10.1186/s40164-023-00407-0. PMID: 37221625; PMCID: PMC10204220.