






eIF4E重編程可變剪接
異常剪接通常歸因于剪接因子(SF)突變,并與包括急性髓系白血病(AML)在內的惡性腫瘤有關。發現了一種與突變無關的方法,即真核轉錄起始因子eIF4E,也能廣泛地重編程可變剪接(AS)。本文于2023年2月發表在《EMBO JOURNAL》期刊上。
技術路線:

主要實驗結果:
1、eIF4E過表達通過其RNA輸出功能影響剪接因子的蛋白水平
作為研究eIF4E在剪接中可能作用的第一步,作者檢查了先前收集的內源性核eIF4E RIP-seq數據集,該數據集來自侵襲性B細胞淋巴瘤LY1細胞系,其中核eIF4E與約3000個轉錄本相關。作者分析顯示,eIF4E與約100個RNA相關,這些RNA編碼參與剪接調節的因子,其中53個RNA編碼主要剪接體的組分,包括SF3B1和U2AF1,這些因子在AML中經常突變。有趣的是,STRING分析揭示所有主要的剪接復合體的顯著富集,根據剪接組的不同,FDR范圍從0.0016到2.19×10-49不等(圖1A)。這個結果促使作者分析內源性eIF4E是否可以調節剪接因子的產生,從而調節剪接。
為探究潛在的eIF4E剪接軸,作者基于以下幾個原因優先使用U2OS細胞進行研究。首先,與上述淋巴瘤細胞不同,U2OS細胞具有內源性eIF4E水平,這與健康的志愿者細胞相似,使作者能夠測量eIF4E過表達的影響。其次,在細胞核中觀察到內源性和過表達的eIF4E,這與剪接和細胞質中的潛在作用一致(圖1B-H)。第三,eIF4E在這些細胞的capping、CPA、核RNA輸出和翻譯中發揮著公認的cap - dependent作用。最后,淋巴瘤和U2OS細胞之間這些相互作用的保守性表明這是eIF4E的一般功能,而不是淋巴瘤特有的現象。考慮到這些,作者生成了3個穩定的2FLAG-eIF4E過表達或vector對照細胞株。首先確定eIF4E是否調節SFs的水平。WB顯示,在2FLAG-eIF4E細胞中,SF3B1、U2AF1、U2AF2、PRPF6、PRPF8、PRPF19、PRPF31、SNRNP200蛋白水平較Vector對照組升高約2 ~ 3倍(圖1C)。CyclinD1和Mcl1作為陽性對照,而b-actin作為陰性對照,因為它們不是eIF4E的靶標(圖1C)。與過表達相反,使用CRISPR-eIF4E或CRISPR-Ctrl U2OS細胞系檢測結果顯示內源性eIF4E敲低會降低這些SFs的水平(圖1D)。這些發現表明內源性eIF4E影響剪接機制的產生,因此,eIF4E對SFs的影響并不局限于以eIF4E升高為特征的環境。總之,eIF4E依賴的方式可以驅動SFs的產生。eIF4E靶向的SFs存在于每個主要的剪接體復合體中(圖1A和1E)。
接下來探究eIF4E是否通過核輸出的機制影響SFs蛋白的產生。作者從核裂解物中進行eIF4ERIP以確定eIF4E是否與U2OS細胞中SFs編碼的RNA結合。在IP過程中,核裂解物與甲醛交聯以防止重配,并通過RT-qPCR監測RNA。觀察到含有SF3B1,SNRNP200,PRPF6,PRPF8,PRPF31,U2AF1,U2AF2的內源性核eIF4ERIP相比于input,被富集了約2到4倍(圖1F)。鑒于這種相互作用,作者研究了這些SFs編碼RNA是否為eIF4E依賴核輸出的靶標。為此,作者使用上述三種不同的細胞系,通過RT-qPCR檢測2FLAG-eIF4E細胞中相對于Vector對照的細胞核和細胞質部分的RNA水平,發現它們都在eIF4E過表達后胞質/核比率增加了~2倍,表明eIF4E過表達增加了它們的核輸出(圖1G)。分級質控采用半定量PCR檢測U6snRNA和tRNAMet,其分別表示細胞核和細胞質部分(圖1G)。總之,這些發現表明,eIF4E增強SFs編碼RNA核輸出,這為eIF4E介導的SFs蛋白產生的升高提供了一種機制。
除了通過核RNA輸出來驅動SFs的產生外,eIF4E還可能間接驅動轉錄和/或改變SFs編碼RNA的轉錄穩定性,作為驅動其蛋白質產生的平行手段。然而,作者注意到文獻中典型的eIF4E靶點,eIF4E過表達不調節任何RNA的總水平(圖1H)。因此,eIF4E不影響這些SF編碼轉錄本的穩態轉錄或RNA穩定性。
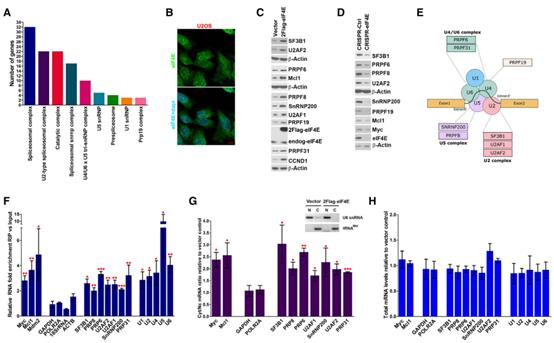
圖1eIF4E通過其核運輸活性改變SF景觀
為分析eIF4E依賴的RNA輸出與其翻譯功能對SF蛋白產生的相對貢獻,作者使用了一個功能分離突變體。eIF4ES53A突變體具有完全結構,結合m7Gcap,促進eIF4E依賴的翻譯和野生型eIF4E;然而,它不能促進依賴eIF4E的核內RNA輸出。作者比較了2FLAG-eIF4E、S53A突變體和Vector對照的SFs蛋白產量。檢測了每個細胞系的3個穩定克隆,并注意到野生型和S53AeIF4E表達水平相似(圖2A)。正如預期的那樣,作為eIF4E-RNA輸出的既定目標,如CCND1,S53A突變體不能刺激細胞cyclinD1蛋白的產生,其水平與Vector對照組相似,而在野生型eIF4E過表達的細胞中,其水平如預期的那樣升高。總之,對于所測試的SFs,S53A突變體沒有像野生型eIF4E那樣促進蛋白表達,這證實了eIF4E在SFs蛋白升高中具有核輸出作用。
為更深入地研究一些SFs在RNA輸出和翻譯水平上共同調控的可能性,作者使用多聚體裝載來直接測量翻譯效率,即每個轉錄本裝載的核糖體數量(圖2B)。如預期的,2FLAG-eIF4E和Vector細胞的整體多核糖體特征是不可區分的(圖2B)。在較重的餾分中發現的轉錄本,每個轉錄本有更多的核糖體,因此比在較輕的餾分中發現的轉錄本翻譯效率更高。用RT-qPCR比較了2FLAG-eIF4E和Vector U2OS細胞系中單體、輕質和重質多核糖體部分的SF編碼RNA含量(圖2B)。預計在翻譯水平上受到影響的RNA在重核糖體部分會增加,同時在輕核糖體和單體部分會減少。發現ACTB mRNAs沒有因為eIF4E的表達而發生改變,而陽性對照MYC和VEGF則如預期的那樣轉移到較重的核糖體中。PRP31、U2AF1、SNRNP200或PRPF6轉錄物的核糖體負荷沒有變化。然而,PRPF8、SF3B1和U2AF2 mRNAs的特點是eIF4E依賴性地轉移到重多核糖體,并在輕核糖體和單體部分減少(圖2B,中間部分)。與S53A突變體觀察到的蛋白水平一致,觀察到這里的大多數eIF4E目標是核輸出目標;然而,少數目標在輸出和翻譯水平上都對eIF4E敏感,使得它們對eIF4E水平更加敏感。
綜上所述,eIF4E與許多SF編碼RNA相互作用并促進其核出口。此外,eIF4E并沒有調節這些轉錄物的穩態轉錄或穩定性。對S53A突變體的研究強烈支持eIF4E的核RNA輸出功能在驅動SF蛋白生產中的主要作用,但對一些SFs來說,也有翻譯的貢獻,使這些后一種RNA對eIF4E失調更加敏感。
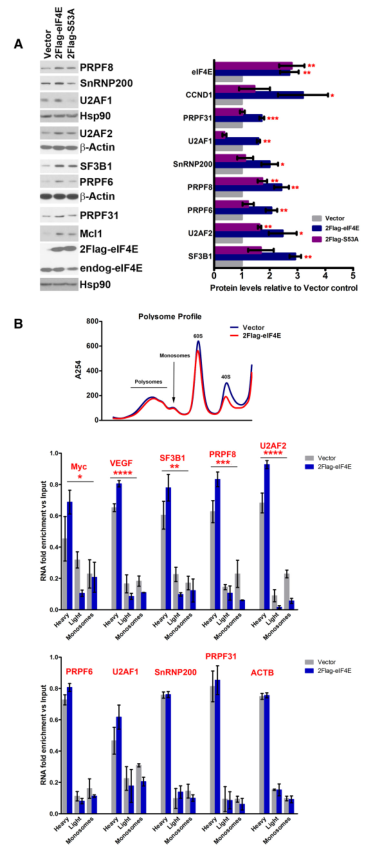
圖2分析S53AeIF4E突變體對其改變SF景觀能力的影響及其在翻譯中的作用
2、eIF4E在AML細胞系和原發性AML標本中驅動SFs的產生
為從遺傳學角度剖析eIF4E在AML背景下的作用,并與U2OS進行比較,以確定這些eIF4E機制是否在不同的細胞類型中是保守的,構建了eIF4E或Vector NOMO-1穩定細胞系。NOMO-1 eIF4E細胞產生的eIF4E蛋白水平和定位與高eIF4E AML標本相似(圖3A)。相比于對照組,這些SFs蛋白水平在NOMO-1 eIF4E細胞中上調(圖3A)。而使用eIF4E inhibitor ribavirin的效果則相反(圖3B)。因此,eIF4E對SFs的調控也適用于AML細胞。
接下來,研究原發性AML標本中eIF4E狀態是否與SFs水平相關。如圖3C所示,結果和AML細胞類似,相比于健康或正常組,eIF4E高表達的AML患者組SFs蛋白水平顯著升高(圖3C)。因此,這些SF的升高并不是AML患者標本的普遍特征,而是與eIF4E水平相關,這與在U2OS和NOMO-1細胞中的研究一致。
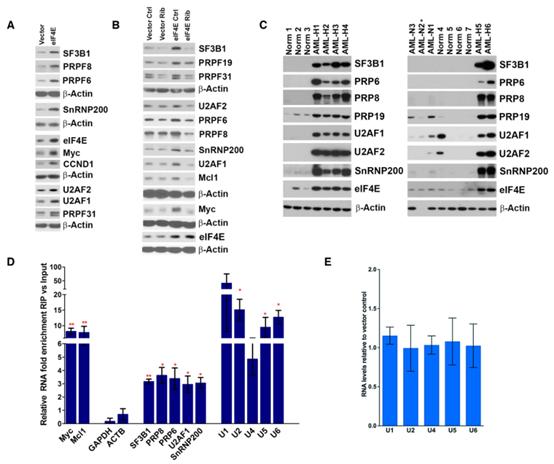
圖3eIF4E在AML中重編程SF景觀
隨后檢測eIF4E是否與SFs相互作用,為eIF4E可能影響剪接的提供額外手段。在RIPs之前,核裂解物用甲醛交聯,以防止處理過程中的重新組合。觀察到內源性eIF4E與U1、U2、U4、U5和U6snRNAs免疫共沉淀,相比于input,在U2OS細胞(圖1F)以及NOMO-1細胞(圖3D)的核裂解物中富集了約3至10倍。內源性eIF4E與陰性對照(GAPDH和ACTB)無關。此外,在U2OS或NOMO-1細胞中eIF4E過表達后,UsnRNAs的水平沒有改變,這表明eIF4E沒有導致更多剪接體的產生(圖1H和3E)。鑒于UsnRNAs在剪接體中發揮結構和催化作用,作者研究了eIF4E是否與剪接體的蛋白質組分物理相關,結果發現內源性eIF4E與U2OS(圖4A)和NOMO-1細胞(圖4B)中每個主要剪接體復合體的蛋白組分發生免疫沉淀,但不沉淀陰性對照。因此,正如在eIF4E依賴capping和CPA中觀察到的那樣,eIF4E促進SFs的產生可能是為了增加其與它們的蛋白質形式物理相互作用的能力。總之,eIF4E與幾個主要剪接體的SF和UsnRNAs結合,并在AML和U2OS細胞環境中提高了SF蛋白水平。因此,eIF4E被定位為在不同的細胞環境中調節剪接,這表明這可能是eIF4E的一個廣泛適用的特性。
4、eIF4E與剪接體的相互作用是RNA和m7Gcap敏感的
如上文所述進行免疫沉淀,只是由于RNAse步驟,U2OS細胞的核裂解物沒有甲醛交聯,然而,即使在沒有交聯的情況下,在未處理的對照組中,SFs和eIF4E之間也觀察到相同的相互作用(圖4A vs 4C)。觀察到SF3B1,U2AF1,U2AF2,PRPF6和PRPF8都存在于未處理的對照中,并且這種相互作用在RNAse處理后減少了約5倍(圖4C)。分離對照表明,作為核標志物的Lamin在這些核裂解物中富集,而MEK1作為細胞質標志物缺乏,此外,在處理或未處理的樣本中,eIF4E均未與Lamin免疫沉淀(圖4C)。因此,eIF4E-SF相互作用需要完整的RNA。
考慮到RNAse處理同時針對底物m7Gcap的pre-mRNAs和UsnRNAs,作者研究了用m7Gcap的類似物處理的影響,這將破壞m7Gcap的mRNA相互作用而不影響UsnRNA介導的相互作用。進行eIF4ERIPs作為m7GpppG或陰性GpppG對照的功能,并通過WB監測SF的關聯(圖4D)。關于RNAse處理,核裂解物沒有甲醛交聯,eIF4E在m7GpppG或GpppG處理的樣品中都能免疫沉淀自身(圖4D)。此外,eIF4E-4EBP1的相互作用沒有受到m7GpppG或GpppG的影響,eIF4E-IP與陰性對照H2B也沒有影響。相對于GpppG對照組,eIF4E與PRPF6、U2AF3和SF3B1之間的相互作用被m7GpppG處理減少了2-5倍(圖4D);此外,HuR/ ELAVL1-eIF4E復合物在m7GpppG處理下也減少了約5倍(圖4D)。鑒于eIF4E通過m7Gcap相互作用與大多數RNA相關,這些研究表明,eIF4E和SF的相互作用是由底物m7GcapRNA介導和/或穩定的。這些結果表明,eIF4E招募了選定的m7Gcap的RNA到剪接體,并且/或者在那里穩定了它們的相互作用。

圖4eIF4E以RNA和帽依賴的方式與剪接機制發生物理相互作用
5、單獨過表達eIF4E足以改變剪接程序
基于上述發現,作者監測了eIF4E對U2OS細胞整體水平剪接的影響。使用rMATS,觀察到約760個轉錄物的約890個剪接事件被改變(FDR-adjusted P-value < 0.15; inclusion differences of > 0.05 or < 0.05),使用更嚴格的截斷值(Pvalue< 0.1, inclusion level differences of > 0.1 or < 0.1),觀察到493個轉錄物的555個剪接事件(圖5B)。在RNA-Seq實驗中,共檢測到TPM大于10的5738個被注釋的轉錄物,表明約15%的被注釋的轉錄物是以eIF4E-dependent的方式進行差異剪接的。SE是最頻繁的事件(約55%),其次是MXE事件(約30%)(圖5A)。聚類熱圖顯示所有類型的剪接事件僅基于eIF4E水平被分離(圖5B)。GO分析顯示,這些轉錄本參與的GO條目包括細胞周期、膜運輸、DNA修復和微管細胞骨架組織。PPI富集分析與GO一致,排名靠前的是細胞周期,RNA代謝和膜運輸(圖5C)。這些結果表明單獨過表達導致AS景觀改變。
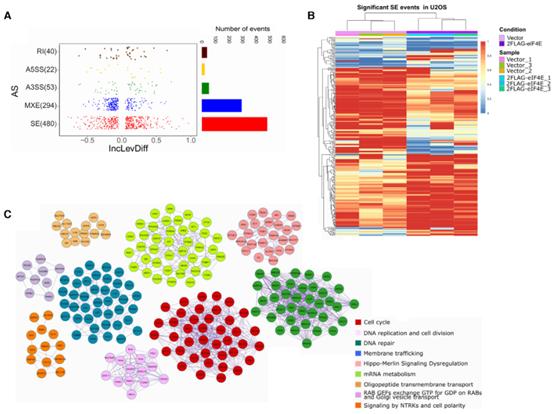
6、在AML中eIF4E水平與差異剪接程序相關
接下來研究了eIF4E依賴性剪接在AML中的相關性,因為在AML中用利ribavirin靶向eIF4E提供了臨床益處,包括在早期臨床試驗中AML患者中的一部分得到緩解。第一步,作者分析了來自437份AML患者樣本和17份來源于人臍帶血的正常CD34+細胞樣本的RNA-Seq數據(圖6A-C)。通過RNA-Seq數據對樣本進行了預篩選,以確保樣本不攜帶AML中報告的主要突變,即U2AF1、SRSF2或SF3B1的突變,使用正常CD34+細胞作為eIF4E水平基準,將AML標本分為高eIF4E組和正常eIF4E組(圖6A-B)。在rMATS分析中,選擇10個eIF4E水平最高的AML標本和11個eIF4E水平正常的標本,即與正常CD34+細胞中的水平重疊或低于其水平(圖6A-C)。相應的臨床數據顯示,與11名eIF4E水平最低的AML患者(1396天,圖6C)相比,10名高eIF4E的AML患者的總生存期急劇下降(中位生存期136天,圖6C)。用rMATS比較這些高eIF4E和正常eIF4E標本的剪接譜,觀察到~1,600個差異剪接事件,影響到~1,500個RNA(FDR-adjusted Pvalue< 0.1, absolute value of inclusion level differences of > 0.1),如果降低閾值((absolute inclusion value difference of > 0.05 and FDR-adjusted Pvalue< 0.15),則有~12,000個差異剪接事件影響~4,600個轉錄物(圖6D)。對這些AS事件進行無監督聚類分析,發現AML患者eIF4E表達狀態與MXE事件密切相關,在監測MXE事件時,20/21例AML標本的eIF4E水平是聚集的基礎(圖6E)。與U2OS細胞相似,觀察到eIF4E依賴的MXE靶點的前GO富集項與U2OS數據集中的高度相似:細胞周期,DNA修復,膜運輸和RNA代謝,PPI富集于有絲分裂中心體、膜運輸和notch信號通路(圖6F)。U2OS和AML數據集中所有eIF4E依賴的剪接靶點的交集獲得了一組核心的約450個常見RNA靶點(圖7A)。類似的,GO和PPI分析顯示它們也富集到細胞周期,DNA修復,膜運輸,染色質修飾,適應性免疫系統,MYC激活等(圖7B)。
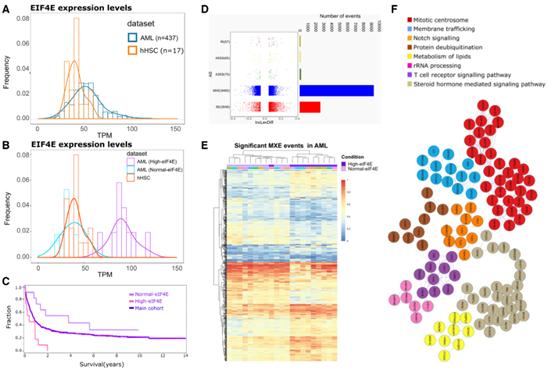
圖6 AML中eIF4E依賴的AS事件
7、eIF4E AS靶向的一個子集與核eIF4E發生物理相互作用
作者使用RT–qPCR驗證了圖7A中核心拼接目標中的幾個eIF4E誘導的可變剪接事件(圖7C)。作者調查了eIF4E是否在剪接體之前(或與剪接體一起)被招募到pre-mRNA上?或者這些eIF4E-轉錄物的聯系是否是剪接產物特異性的。為解決這個問題,使用甲醛交聯進行核eIF4ERIPs并使用特異性引物RT-qPCR分析RNA含量。結果與用底物和產物RNA的引物獲得的RNA的水平進行了比較,后者作為正常化對照。觀察到,eIF4ERIPs與尚未經歷目標剪接事件("未剪接")的轉錄本,以及那些已經經歷了剪接事件("已剪接")的IL1B、IL4R、IKBKB、FAAP20和MAPK8IP3富集(圖7D)。這些發現表明eIF4E在至少某些剪接靶點的剪接中起直接作用。此外,這些研究首次表明eIF4E與pre-mRNA結合,這對eIF4E的功能具有重要意義。
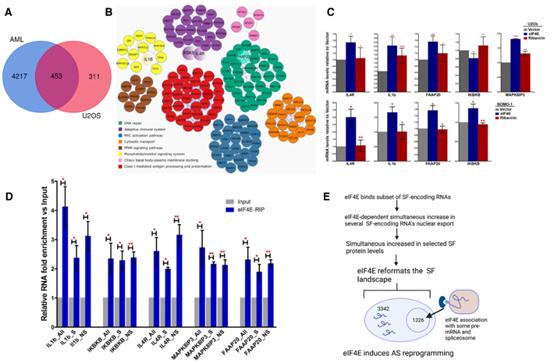
圖7識別和驗證核心剪接網絡以及與pre-mRNAs AS靶點的物理相互作用
總之,本研究證實,核eIF4E與SF編碼的RNA有物理聯系,并驅動它們的核輸出,這些轉錄物的核輸出增強反過來又增加了它們對翻譯機器的可用性,并提高了SF蛋白的水平,并且eIF4E在整體水平重建了細胞SFs(圖7E)。
參考文獻:
Ghram M, Morris G, Culjkovic-Kraljacic B, Mars JC, Gendron P, Skrabanek L, Revuelta MV, Cerchietti L, Guzman ML, Borden KLB. The eukaryotic translation initiation factor eIF4E reprograms alternative splicing. EMBO J. 2023 Feb 27:e110496. doi: 10.15252/embj.2021110496. Epub ahead of print. PMID: 36843541.