






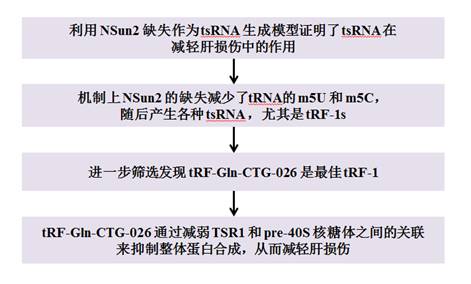
tRF-Gln-CTG-026通過減輕整體蛋白合成來改善肝損傷
肝臟執行各種生物學功能以維持體內平衡,如糖原儲存、營養代謝、膽汁分泌和蛋白質合成。當發生肝損傷時,肝細胞會經歷巨大的損傷應激,導致出血性壞死、肝細胞凋亡、肝脂肪變性和肝臟炎癥等。由急性肝衰竭、肝硬化、癌癥及其他原因引起的肝損傷是全世界眾多死亡的重要原因。肝移植幾乎是促進生存的唯一治療方法,卻受副作用和供體短缺的限制。肝臟可以有效地將小RNA輸送到其中,小RNA療法可能為肝損傷開辟一條新途徑。tsRNA(tRNA衍生的小RNA)參與應激反應,并與損傷有廣泛的關聯。盡管tsRNA在調節應激反應和損傷方面發揮重要作用,但由于缺乏以治療為重點的研究,tsRNA尚未被開發為減輕疾病的小治療RNA。該研究發表于《Signal Transduction and Targeted Therapy》,IF:38.104。
技術路線:
主要研究結果:
1. 建立NSun2缺失作為tsRNA生成模型以減輕體外損傷
作者首先在小鼠中誘導肝損傷,以驗證tsRNA是否參與肝損傷和修復(圖1a)。作者發現在肝損傷小鼠中,tsRNA的數量增加(圖1b),說明tsRNA的產生與肝損傷有關。為了進一步研究tsRNA與肝損傷的關系,有必要建立tsRNA生成模型。作者首先在體外細胞實驗中定義中高、低損傷應激的標準。對H2O2HL-7702和AML12細胞進行劑量梯度實驗。由此,作者將10μM H2O2應激定義為低H2O2應激或增殖應激,不會導致細胞死亡,但會抑制細胞生長;將HL-7702的500μM H2O2應激和AML12的100μM H2O2應激定義為高H2O2應激、致死應激或生存應激。通過篩選HL-2細胞系中的三種tRNA修飾酶(NSun1,Mettl2和Dnmt7702),在低損傷應激下,與ME-KD(Mettl2敲低)和DN-KD(Dnmt1敲低)相比,NSun2敲低(NS-KD)導致增殖增加(圖1c,e);此外,在高應激下,NSun2敲低提高了細胞存活率(圖1d,e)。為了進一步探索NS-KD在細胞損傷中的作用,作者使用生長曲線實驗測量了在低H2O2應激下連續5天的細胞增殖。結果表明,在H2O2處理下,NS-KD促進細胞增殖(圖1f)。相比之下,作者應用了不同的高強度應激誘導劑(CCl4、對乙酰氨基酚和油酸混合物)并測量了細胞存活率。結果表明,NS-KD改善了細胞在應激損傷下的存活率(圖1g,i)。正如預期的那樣,NS-KD僅在應激下起作用。在基礎條件下,NS-KD不影響細胞增殖(圖1h,i)。總之,這些結果表明肝損傷與tsRNA的產生有關,NS-KD促進了體外應激下的細胞增殖和存活,但不改變細胞遷移。
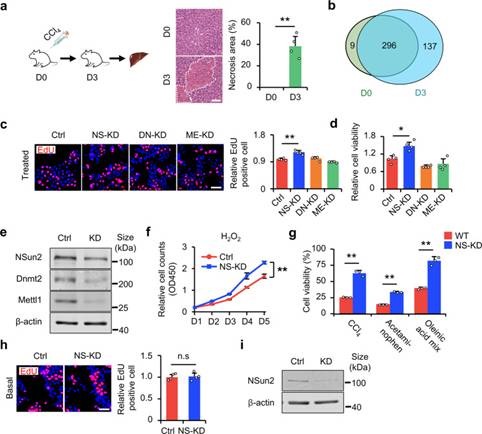
圖1 NS-KD減輕細胞損傷
2. NS-KO減輕肝臟損傷
如上所述,NSun2的缺失是一個合適的研究tsRNA模型,可以提高細胞在應激下的增殖和生存。因此,作者通過CRISPR / Cas9介導的NSun2基因第2外顯子中56 bp缺失產生了NS-KO小鼠(圖2a)。在功能上,作者首先驗證了NSun2缺失突變體是否對肝損傷的應激有反應。在沒有應激損傷的情況下,NS-KO沒有引起肝臟形態和病理生理學的異常(圖2b),這表明NS-KO和由此產生的tsRNA在沒有應激的情況下都沒有發揮作用。通過腹腔注射CCl4建立短期或長期肝損傷模型(圖2c),作者證明了NS-KO對應激損傷有反應,并減輕了肝損傷。在CCl4單次注射后第3天,肝臟出現壞死和炎癥;第27天,大鼠出現肝纖維化。NS-KO小鼠的壞死程度低于野生型(WT)小鼠(圖2d)。檢測單次或重復注射CCl4后的血液生化指標。丙氨酸轉氨酶、天冬氨酸轉氨酶和堿性磷酸酶在CCl4單次注射后第3天達到最大值,肝細胞損傷嚴重,在CCl4重復注射后第27天降至正常水平。NS-KO小鼠的所有三項指標均低于WT小鼠,這意味著NS-KO小鼠的肝損傷輕微(圖2e,f)。為了研究NSun2缺失如何改善肝損傷,作者通過BrdU摻入和Ki67染色檢測肝細胞增殖。與WT小鼠相比,作者發現NS-KO小鼠在肝損傷第3天增殖的肝細胞數量顯著增加(圖2g)。作者還使用原位切口末端標記法檢測了NSun2缺失如何影響細胞死亡。結果提示肝細胞凋亡在肝損傷后第3天達到峰值,然后逐漸下降。與WT小鼠相比,NS-KO小鼠中檢測到較少的凋亡肝細胞,這意味著NSun2的缺失抵抗了細胞凋亡(圖2h)。在應激條件下,促增殖、促生存和抗炎相關基因的表達增加,而抗增殖、抗存活和促炎癥相關基因的表達被抑制(圖2i)。總之,作者觀察到在短期和長期肝損傷方面NS-KO小鼠的肝損傷均比WT小鼠輕,這表明NSun2缺失可減輕體內肝損傷。
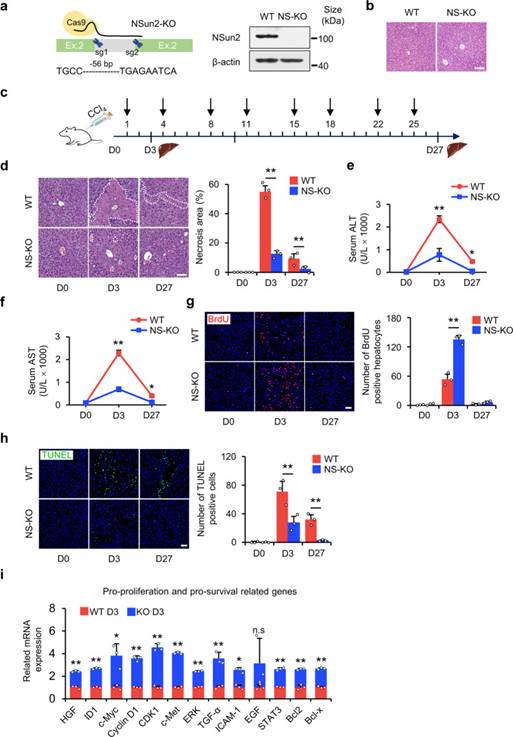
圖2 NS-KO改善體內肝臟壞死、再生和存活
3. NS-KO衍生的小RNA可促進細胞在應激下的增殖和存活
作者假設,NS-KO通過NS-KO衍生的小RNA預防肝損傷。為了驗證這一假設,作者首先在HL-7702細胞系中敲低NSun2基因,將無酶活性的突變型NSun2(K190M或C271A)轉染到HL-7702細胞系中,并對細胞進行細胞毒性刺激。EdU摻入實驗和細胞增殖實驗表明,只有WT NSun2抑制了NSun2敲低引起的細胞增殖促進(圖3a,b,d),而不是NSun2突變體(K190M或C271A)。同樣,只有WT NSun2逆轉了NS-KD誘導的細胞活力增加;NSun2突變體則沒有(圖3c,d)。這些結果表明,NSun2的缺失主要通過tRNA甲基轉移酶活性的缺失促進細胞在應激下的增殖和存活。當NSun2失去甲基轉移酶活性時,tRNA修飾不可避免地減少。在NS-KO細胞中,tRNA表達降低(圖3e)。最顯著的變異是tRNA m5U和前面描述的M5C.13兩者在NS-KO樣品中顯著降低(圖3f,g)。
上述結果表明,NS-KO在減少tRNA m5U和m5C修飾的同時,失去了其甲基轉移酶活性。然后作者研究了NS-KO衍生的小RNA是否足以改善細胞在應激下的增殖和存活。作者從WT和NS-KO小鼠中分離出14-50 nt和50-100 nt的肝RNA片段(作為陰性對照)(圖3h)。由于tsRNA的大小總是小于50 nt,作者認為14-50 nt的RNA是含tsRNA的RNA,50-100 nt的RNA是非tsRNA和陰性對照。作者首先檢測了14-50 nt RNA和50-100 nt RNA 在低H2O2應激下的增殖差異。EdU摻入實驗顯示,與WT小鼠的片段相比,NS-KO小鼠的14-50 nt的RNA片段轉染增加了細胞增殖(圖3i)。同樣,CCK-8檢測表明,來自NS-KO小鼠的14-50 nt RNA片段增加了損傷后的細胞存活率,而來自WT小鼠的14-50 nt RNA片段和來自兩種小鼠的50-100 nt RNA片段(圖3j)。因此,NS-KO衍生的14-50 nt RNA片段在促進損傷后的細胞增殖和存活中發揮重要作用。
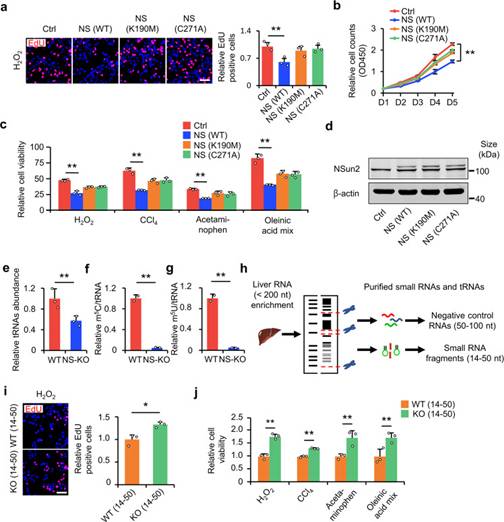
圖3 NS-KO來源的tsRNA可減輕肝損傷
4. tRF-1是NS-KO的重要產物
為了全面分析tsRNA的代謝特性,分析WT和NS-KO小鼠在不同應激損傷后tsRNA的差異,作者首先確定了NSKO對miRNA的數量沒有明顯改變(WT52.96%,NS-KO 53.12%)。通過tRNA衍生的小RNA測序(tsRNA-seq),作者鑒定了tRFdb數據庫中記錄的841個未記錄的tsRNA和104個已知的tsRNA(圖4a)。然后,作者通過比較來自WT和NS-KO小鼠的樣品來分析tsRNA表達,并鑒定出366個上調和267個下調的tsRNA,表明NS-KO引起的tsRNA上調多于下調。有趣的是,NS-KO小鼠中的tsRNA在整個損傷過程中保持相對較高的水平,而WT小鼠的tsRNA在D3達到峰值(圖4b)。因此,作者假設在整個CCl重復注射期間,第3天是肝細胞增殖和tsRNA生成最活躍的時間(圖2g,4b)。這些結果表明,NSun2缺失增加了NS-KO小鼠的tsRNA,并預留了足夠的tsRNA來應對應激損傷,但在WT小鼠中,tsRNA只能在WT D3產生。
在進一步分析了WT和NS-KO小鼠在肝損傷不同階段的tsRNA景觀后,作者發現與其他tsRNA相比,NS-KO小鼠中的tRF-1s突然增加。在肝損傷的各個時間點,NS-KO組的tRF-1s明顯多于WT組,但WT組僅在第3天增加(圖4c,d)。對tRF-1s的詳細分析發現,tRF-1s的長度主要在14-23 nt之間(圖4d)。一般來說,NSun2表達缺失后,肝臟主要產生14-23nt的tRF-1。作者還檢測了tRF-1s的表達,發現其在NS-KO小鼠中的表達高于WT小鼠,而其他類型的tsRNA在NS-KO小鼠中的表達低于WT小鼠(圖4e)。同時,基于WT和KO以及WT D0和WT D3之間的差異,作者確定了68個可能起作用的tRF-1。因此,作者確定了tRF-1在NS-KO中增加,分析了tRF-1的分布,并確定了68個NS-KO衍生的tRF-1是預防肝損傷和響應應激損傷的候選tsRNA。
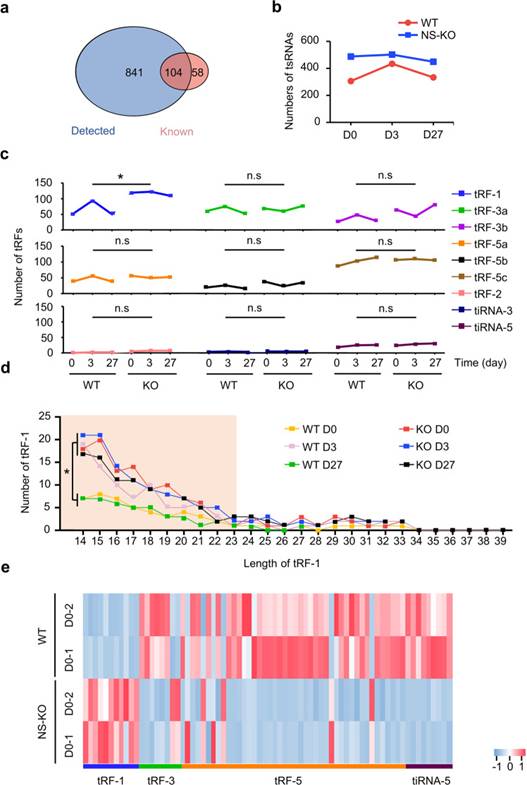
圖4 tsRNA-seq顯示了tRF-1在WT和NS-KO小鼠中的特征
5. 篩選的tRF-1在體外和體內挽救肝損傷
如上所述,肝損傷的副產物主要是tRF-1。為了闡明tRF-1s是否可以作為肝損傷的潛在治療方法,作者人工合成了圖4所示的68個tRF-1。作者用硫代磷酸酯(PS)、2'-O-甲基(2'-OMe)和膽固醇修飾了tRF-1s,以提高其在體內的穩定性和轉染效率(圖5a)。將人工合成的tRF-1轉染HL-7702細胞和人原代肝細胞(PHH),并在體外和體外驗證其功能。EdU摻入試驗顯示合成的tRF-1s促進細胞增殖(圖5b)。CCK-8實驗顯示過表達tRF-1s后細胞活力提高(圖5c)。在PHH實驗中,作者也得到了一致的結果(圖5d,e)。為了驗證修飾的合成tRF-1s在體內的功能,作者將tRF-1s靜脈注射到肝損傷小鼠體內。注射68個tRF-1的結果顯示損傷減少,肝功能改善(圖5F-i)。這些結果表明,68個tRF-1s在體外和體內加速了受損肝細胞的恢復。綜上所述,作者發現作者篩選出的tG026在體內減輕了肝損傷(圖5J,K),而在NS-KO后tG026的表達增加(圖5l,m)。總之,通過篩選,作者確定了tG026可以減輕肝損傷。

圖5 篩選的tG026促進損傷后的細胞增殖和存活
6. tRF-Gln-CTG-026通過調節核糖體組裝減少全球整體蛋白質合成
為了探索tG026的潛在機制,作者進行了RNApull-down-LC-MS/MS測定,以篩選與tG026相互作用的蛋白質(圖6a)。在尋找與HL-7702和AML12細胞系的tG026共同相互作用的蛋白后,作者選擇了參與Pre-rRNA加工蛋白TSR1同源物進行后續實驗(圖6b)。作者推測tG026可能調節核糖體。通過RNApull-down實驗,作者在兩個細胞系中驗證了TSR1和tG026之間的相互作用(圖6c,d)。為了確定tG026如何調節核糖體組裝,作者在HL-7702和AML12細胞中過表達tG026。結果表明,tG026不改變18S rRNA的表達,但抑制了TSR1和18S rRNA之間的關聯。這表明tG026抑制TSR1與核糖體之間的相互作用,因為18S rRNA是核糖體復合物的重要組成部分(圖6e,f)。最后,tG026抑制GPS(圖6g,h)。綜上所述,NSun2缺失會導致tG026的產生。tG026通過抑制TSR1與18S rRNA之間的關聯來緩解GPS,以促進肝損傷后的增殖和存活,可以開發為緩解肝損傷的RNA藥物。
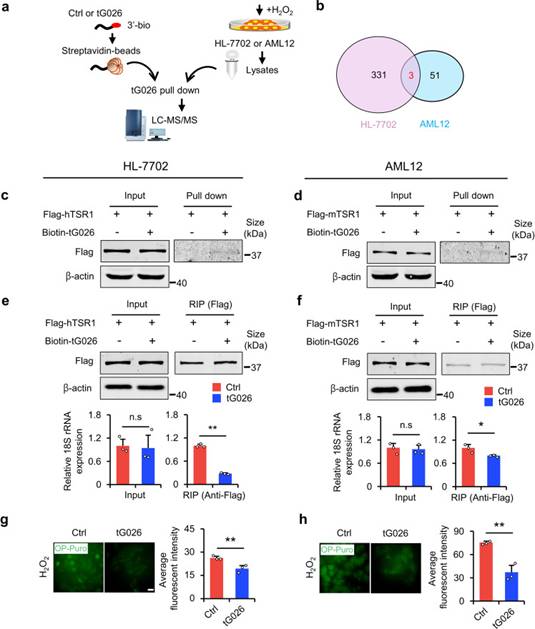
圖6 tG026通過抑制TSR1與18S rRNA之間的相互作用來降低GPS
結論:
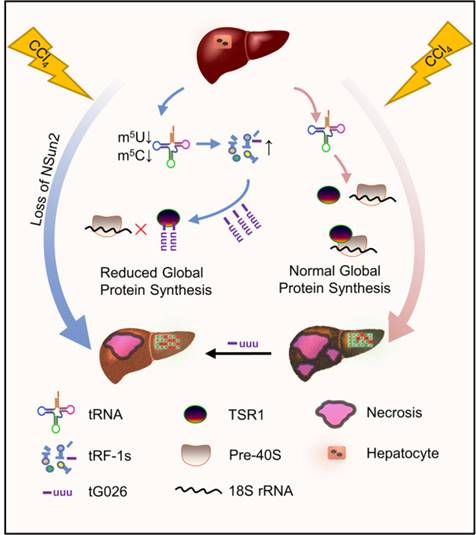
該研究利用調節GPS的tRNA降解產物RNA降低了RNA毒性。相關機制是通過tG026介導的TSR1和18S rRNA之間相互作用的破壞,這是由于tG026與TSR1的相互作用,阻止了TSR1和Pre-40S核糖體之間的關聯。這導致核糖體組裝程序的抑制,從而降低GPS。
tG026在肝損傷中的機制模型圖:
參考文獻:
Ying S, Li P, Wang J, et al. tRF-Gln-CTG-026 ameliorates liver injury by alleviating global protein synthesis. Signal Transduct TargetTher. 2023 Apr 3;8(1):144.