






pTINCR微蛋白通過CDC42 SUMO化和激活促進上皮分化,抑制腫瘤生長
人類轉錄組包含數千個小的開放閱讀框(sorf),它們編碼的微蛋白的功能在很大程度上仍未被探索。在本研究中,我們發現TINCR lncRNA編碼pTINCR, pTINCR是一種進化保守的泛素樣蛋白(UBL),在許多上皮細胞中表達,在分化和細胞應激下表達上調。通過功能獲得和功能喪失的研究,我們證明了pTINCR是體外和體內上皮分化的一個關鍵誘導因子。有趣的是,在幾種上皮性癌癥中,TINCR的低表達與較差的預后相關,而pTINCR的過表達降低了患者源性異種移植瘤的惡性程度。在分子水平上,pTINCR通過SUMO相互作用motif (SIM)與SUMO結合,并與CDC42結合,CDC42是一種RhoGTPase,對肌動蛋白細胞骨架重塑和上皮分化至關重要。此外,pTINCR增加CDC42的sumo化并促進其激活,觸發促分化級聯反應。我們的研究結果表明,微蛋白質組是與癌癥相關的細胞身份的新調控源。本文于2022年11月11日發表于Nature Communications(IF=17.694)。
技術路線:

主要研究結果:
(1) pTINCR是由TINCR lncRNA編碼的一種保守微蛋白,在上皮組織中表達
TINCR是角化細胞和其他上皮細胞分化過程中上調的3.7千堿基lncRNA 40 - 42。我們證實了TINCR在人和小鼠皮膚中表達,也在其他上皮細胞中表達(圖1b)。對TINCR轉錄本的保守分析顯示sORF為264 bp(圖1c)。小鼠皮膚核糖體譜分析(RibORF評分≥0.7)顯示,該sORF被翻譯成一種高度保守的具有87個氨基酸的微蛋白,我們將其命名為pTINCR(圖1c, d)。為了確認pTINCR sORF被翻譯成一種穩定的微蛋白,我們在35s -蛋氨酸存在的情況下,使用全長TINCR lncRNA (integrl: ENST00000448587.5)進行了體外翻譯(圖1e, f),并獲得了約12 kDa的肽產物。值得注意的是,突變pTINCR的起始密碼子會損害任何可檢測產物的翻譯(圖1e, f)。
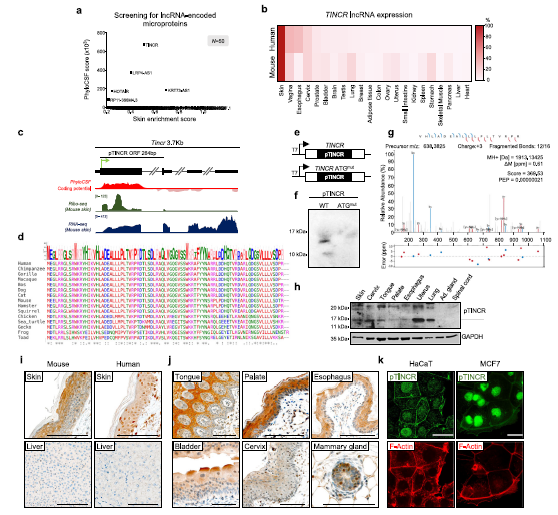
(2) pTINCR促進體外和體內上皮分化
為了表征pTINCR的細胞和分子功能,我們在強力環素誘導的慢病毒載體中克隆了標記HA表位的pTINCR sORF。我們生成了兩種不同的結構,將ha標記放置在微蛋白的c端(pTINCR-HA)或n端(HA-pTINCR)部分。為了最大限度地減少標簽對pTINCR的可能影響,我們在HA和pTINCR sORF之間引入了一個靈活的連接器。此外,為了分離TINCR lncRNA和pTINCR微蛋白的功能,我們還通過突變約20%的pTINCR-HA核苷酸序列,開發了一個合成的ORF (syORF),在生產相同的蛋白質的同時顯著改變了RNA的二級結構。我們在整個手稿中使用syORF來驗證我們的主要發現(見圖圖例)。在U2OS中瞬時表達時檢測到C-和N-末端結構,并顯示出類似的分布,在所有細胞組分中均可檢測到,但最顯著的是在細胞核和細胞間連接處,與蛋白質的內源性定位一致(圖1k)。
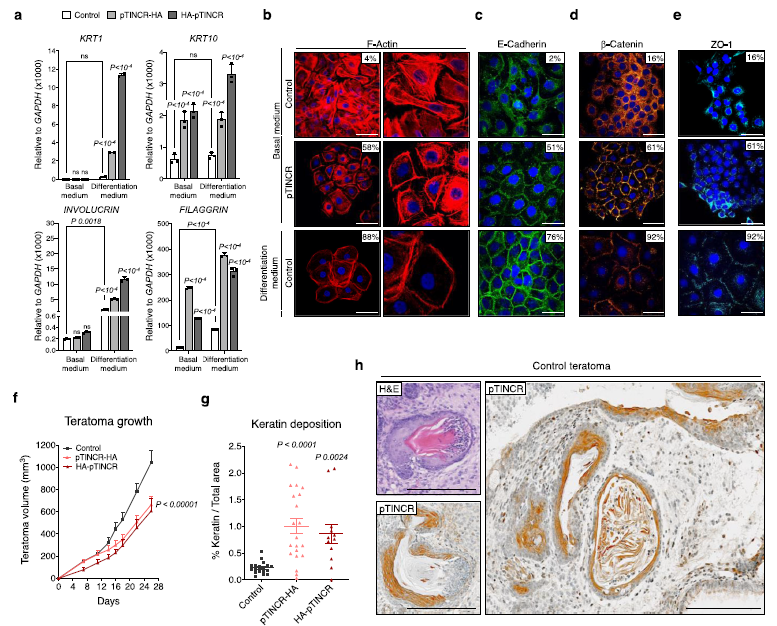
(3) 體外上皮分化需要pTINCR
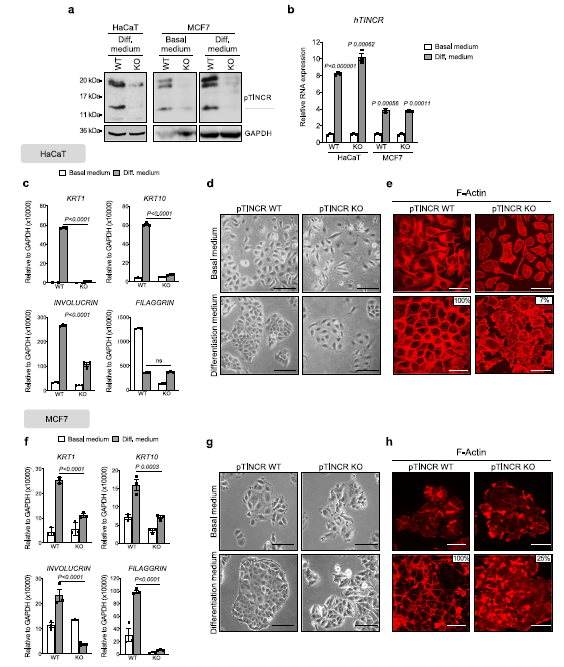
我們使HaCaT和MCF7細胞都受到高鈣的作用,高鈣是一種眾所周知的表皮分化誘導劑50,同時也是其他上皮組織如食管或乳腺的分化誘導劑。WTMCF7細胞(圖4a) O n, p t I n C R p R O e x p R s n n t M cf7細胞(圖4a)。重要的是,我們觀察到在WT和pTINCR-KO細胞中,TINCR lncRNA在鈣誘導的分化過程中有相同程度的上調(圖4b),表明在工程pTINCR-KO細胞中,TINCR lncRNA的調控不受影響。然而,ptincro缺陷細胞無法像WT細胞一樣上調分化標志物(圖4c, f)。此外,pTINCRKO細胞在分化條件下沒有獲得上皮形態(圖4d, g),也沒有重塑其肌動蛋白細胞骨架向皮層傾向(圖4e, h)。這些結果表明,pTINCR蛋白獨立于TINCR lncRNA,在體外實現完全上皮分化是必需的。
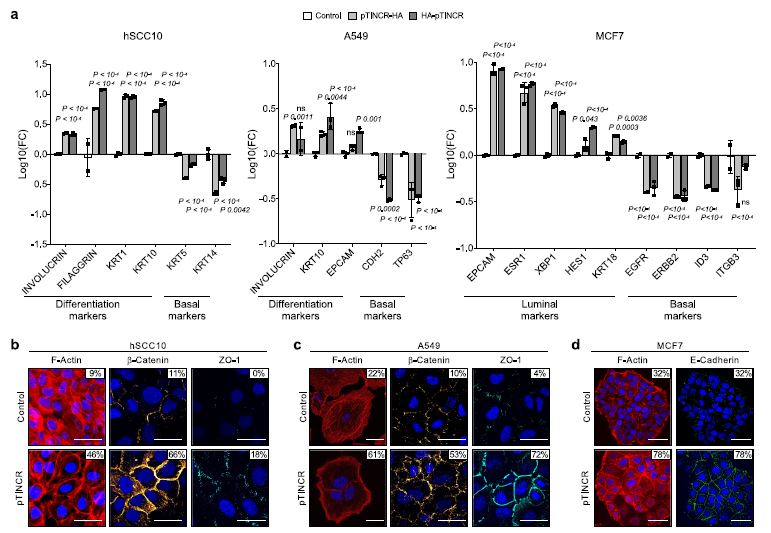
(4) pTINCR觸發上皮分化轉錄程序
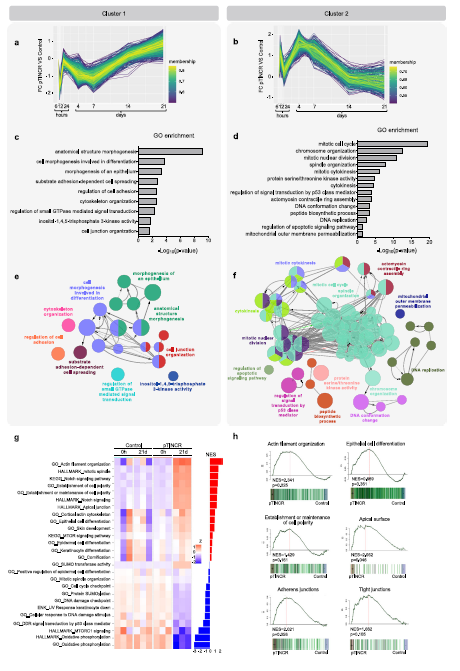
為了證實pTINCR的促分化功能,我們分析了pTINCR過表達誘導的轉錄譜。我們在pTINCR過表達的hSCC10細胞系中進行了廣泛的RNA-seq分析,以評估早期轉錄組變化(6、12和24小時)和長期變化(4、7、14和21天),后者可能反映了細胞特性的變化。首先,我們使用脈沖算法研究了pTINCR過表達驅動的轉錄動力學,脈沖算法是縱向測序實驗的框架,揭示了與time54相關的差異基因表達。我們檢測到6組不同的基因,根據它們的動態表達聚集在一起(圖5a, b)。我們對每個聚類進行了基因本體富集分析(圖5c, d)。有趣的是,其中兩個簇1和簇2是動態對立的簇,都豐富了與細胞骨架相關的基因本體術語。
(5) pTINCR在細胞應激時以p53依賴的方式上調,是損傷誘導分化所必需的
pTINCR過表達21天后,其他上皮特異性的信號(“上皮細胞分化”、“細胞-細胞連接”、“上皮細胞-細胞粘附”、“頂端表面”、“Notch信號”)和表皮相關的信號(“表皮細胞分化”、“皮膚發育”、“角化細胞分化”、“角化”)也會發生變化(圖5g, h)。與pTINCR表達負相關的基因集在與細胞周期、細胞代謝和蛋白質加工等相關的通路中富集(圖5g)。有趣的是,我們發現ptincr下調基因也富集在“Myc靶點”的信號區,這是cSCC中的一種致癌途徑,與腫瘤的分化級別和該惡性腫瘤的臨床預后密切相關44,56,57。我們通過RT-qPCR分析驗證了這些結果。總之,該轉錄組學研究強烈支持我們的功能研究揭示的pTINCR促分化功能。
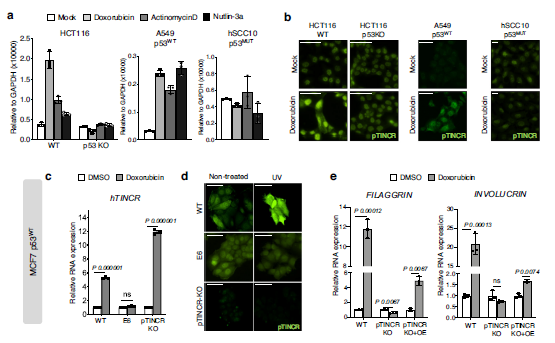
眾所周知,P53是一種腫瘤抑制因子,在50%以上的人類癌癥中都發生了突變。此外,越來越多的證據表明p53也參與胚胎發育和細胞分化58,59。我們之前的研究結果表明,考慮到HaCaT細胞是p53突變體,pTINCR在鈣誘導分化中的作用不需要功能p53。考慮到在上皮組織中,細胞損傷會激活末端分化程序36,37,我們想知道pTINCR的表達是否與損傷時p53的激活有關。首先,我們用幾種p53誘導劑或穩定劑(阿霉素、放線菌素- d或nutlin-3a)處理不同的癌細胞系,并分析TINCR表達和pTINCR水平。我們觀察到,只有p53功能正常(A549或HCT116)的細胞在mRNA和蛋白水平上調了pTINCR,而p53- ko細胞(HCT116 p53KO)或p53突變(hSCC10)沒有表現出相同的反應(圖6a, b)。
這些結果證實了pTINCR在應激中以p53依賴的方式上調。我們分析了已發表的p53 ChIP-seq實驗60,61,我們沒有觀察到p53與pTINCR位點結合,無論是在對照組細胞中還是在使用dna損傷劑處理的細胞中,這表明p53對pTINCR的調控是間接的。接下來,我們使用pTINCR- ko MCF7細胞系研究了pTINCR缺乏對p53依賴性dna損傷誘導分化的影響。MCF7細胞是p53 WT,正如預期的那樣,在pTINCR-WT和pTINCR-KO細胞中,TINCR轉錄水平在損傷后上調(圖6c)。
結論:
總之,我們揭示了pTINCR是一種以前被忽視的UBL-微蛋白,它調節上皮細胞的識別并顯示出腫瘤抑制活性(圖10)。pTINCR作為一種假設lncRNA的生物活性產物的發現,促進了由lncRNA編碼的微蛋白所代表的額外調控水平的想法。對微蛋白質組的探索可以為調控生理和病理過程(如癌癥)提供新的細胞識別調節因子。
參考文獻:
Boix, O., Martinez, M., Vidal, S., Giménez-Alejandre, M., Palenzuela, L., Lorenzo-Sanz, L., Quevedo, L., Moscoso, O., Ruiz-Orera, J., Ximénez-Embún, P., Ciriaco, N., Nuciforo, P., Stephan-Otto Attolini, C., Albà, M. M., Mu?oz, J., Tian, T. V., Varela, I., Vivancos, A., Ramón Y Cajal, S., Mu?oz, P., … Abad, M. (2022). pTINCR microprotein promotes epithelial differentiation and suppresses tumor growth through CDC42 SUMOylation and activation. Nature communications, 13(1), 6840. https://doi.org/10.1038/s41467-022-34529-6.