






LncRNA MILIP促進(jìn)透明細(xì)胞腎細(xì)胞癌的轉(zhuǎn)移
遠(yuǎn)處轉(zhuǎn)移是透明細(xì)胞腎細(xì)胞癌(ccRCC)相關(guān)死亡率的主要原因。然而,涉及ccRCC轉(zhuǎn)移的分子機(jī)制仍有待充分了解。隨著lncRNAs在癌癥發(fā)展、進(jìn)展和治療耐藥性中的作用越來越被重視,導(dǎo)致ccRCC發(fā)病機(jī)制的異常表達(dá)lncRNAs的列表正在迅速擴(kuò)大。與原發(fā)ccRCC組織相比,轉(zhuǎn)移性ccRCC中MILIP表達(dá)上調(diào)。轉(zhuǎn)移性ccRCC細(xì)胞中MILIP表達(dá)增加是由TFAP2C驅(qū)動(dòng)的。MILIP的敲低降低了ccRCC細(xì)胞在體外遷移和侵襲的潛能,并減少了ccRCC在體內(nèi)轉(zhuǎn)移病灶的形成。MILIP對(duì)ccRCC細(xì)胞的影響與EMT標(biāo)志基因表達(dá)的改變有關(guān)。機(jī)制上,MILIP與Snai1 mRNA形成RNA-RNA雙鏈,并與YBX1結(jié)合。這促進(jìn)了YBX1蛋白和Snai1 mRNA之間的關(guān)聯(lián),導(dǎo)致后者的翻譯增加。Snai1反過來在MILIP驅(qū)動(dòng)的ccRCC轉(zhuǎn)移中發(fā)揮重要作用。總之,TFAP2C響應(yīng)的lncRNA MILIP驅(qū)動(dòng)ccRCC轉(zhuǎn)移。因此,靶向MILIP可能是ccRCC治療的潛在途徑。本文于2022年8月發(fā)表于Journal of Experimental & Clinical Cancer Research ( IF=12.658 )上。
技術(shù)路線

結(jié)果
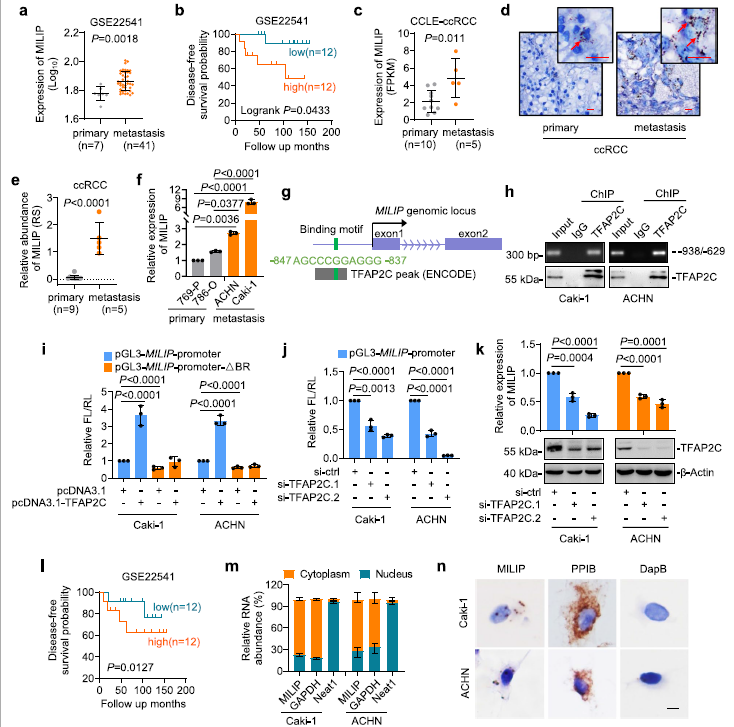
1)TFAP2C驅(qū)動(dòng)轉(zhuǎn)移性ccRCCs中上調(diào)的MILIP表達(dá)
通過查詢ccRCC數(shù)據(jù)集(GSE22541),我們發(fā)現(xiàn),與原發(fā)腫瘤相比,轉(zhuǎn)移病灶來源的ccRCC組織中l(wèi)ncRNA MILIP表達(dá)水平更高(圖1a)。此外,MILIP高表達(dá)與ccRCC患者無病生存期(DFS)較差相關(guān)(圖1b)。對(duì)CCLE的ccRCC細(xì)胞系數(shù)據(jù)集的分析顯示,與原發(fā)腫瘤相比,轉(zhuǎn)移性腫瘤產(chǎn)生的ccRCC細(xì)胞系中MILIP表達(dá)水平更高(圖1c)。我們證實(shí),在FFPE ccRCC樣本隊(duì)列中,與原發(fā)性ccRCC樣本相比,轉(zhuǎn)移性ccRCC中MILIP表達(dá)增加(圖1d, e)。類似地,MILIP表達(dá)水平在轉(zhuǎn)移性細(xì)胞系中更高(cki -1和ACHN) (圖1f)。接著,為了了解在ccRCC細(xì)胞中調(diào)控MILIP表達(dá)的轉(zhuǎn)錄機(jī)制,我們使用生物信息學(xué)查詢其啟動(dòng)子的轉(zhuǎn)錄因子結(jié)合位點(diǎn)。這揭示了位于 MILIP 基因啟動(dòng)子-837/-847 區(qū)域的轉(zhuǎn)錄因子 TFAP2C 的共有結(jié)合基序(圖1g)。來自ENCODE的ChIP-seq數(shù)據(jù)分析顯示,在TFAP2C結(jié)合區(qū)(TFAP2C-BR)有一個(gè)TFAP2C結(jié)合峰(圖1g)。該區(qū)域與內(nèi)源性TFAP2C共同沉淀,是MILIP轉(zhuǎn)錄上調(diào)所必需的(圖1h, i)。此外,TFAP2C的敲除降低了Caki-1和ACHN細(xì)胞中MILIP的轉(zhuǎn)錄活性(圖1j),支持MILIP 通過已識(shí)別的 TFAP2C-BR 被 TFAP2C 轉(zhuǎn)錄激活。為了證實(shí)這一點(diǎn),TFAP2C的敲除降低了內(nèi)源性MILIP水平(圖1k)。此外,與MILIP的高表達(dá)一樣,在GSE22541數(shù)據(jù)集中,TFAP2C的高表達(dá)與ccRCC患者較差的DFS相關(guān)(圖1l)。通過qPCR分析和原位雜交(ISH)分析顯示,MILIP主要位于ccRCC細(xì)胞的細(xì)胞質(zhì)中(圖1m, n)。
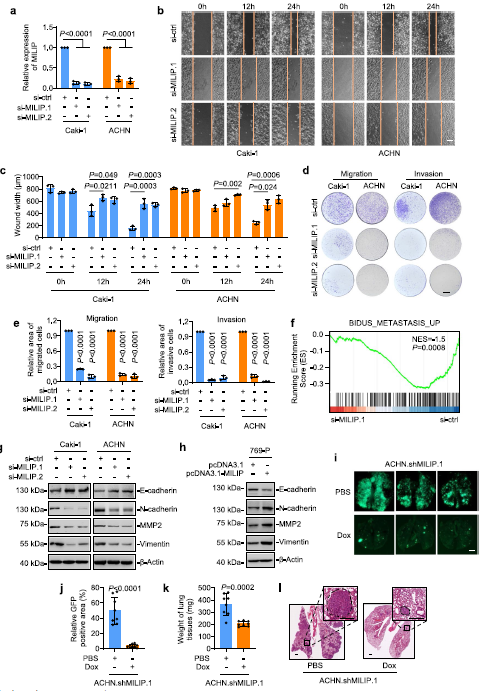
2)MILIP促進(jìn)ccRCC轉(zhuǎn)移
在發(fā)現(xiàn)MILIP在轉(zhuǎn)移性ccRCC中特異性上調(diào)后,我們重點(diǎn)研究了MILIP對(duì)ccRCC細(xì)胞侵襲和轉(zhuǎn)移的潛在作用。MILIP的敲除在劃痕試驗(yàn)中顯著延遲了Caki-1和ACHN細(xì)胞的傷口愈合,并在transwell試驗(yàn)中減少了它們的遷移和侵襲(圖2a-e)。來自Caki-1細(xì)胞的RNA測(cè)序數(shù)據(jù)的GSEA顯示MILIP敲除導(dǎo)致轉(zhuǎn)移途徑基因特征的減少(圖2f)。此外,MILIP的敲低導(dǎo)致上皮標(biāo)記物E-cadherin的上調(diào)以及間充質(zhì)標(biāo)記物N-cadherin、MMP2和波形蛋白的下調(diào),而MILIP過表達(dá)導(dǎo)致E-cadherin的降低,N-cadherin、MMP2和波形蛋白的表達(dá)上調(diào)(圖2g,h)。此外,shMILIP細(xì)胞顯著減少了肺轉(zhuǎn)移病灶的形成(圖2i-l)。總之,這些結(jié)果表明MILIP促進(jìn)ccRCC細(xì)胞的細(xì)胞侵襲和遷移,這與增強(qiáng)EMT有關(guān)。
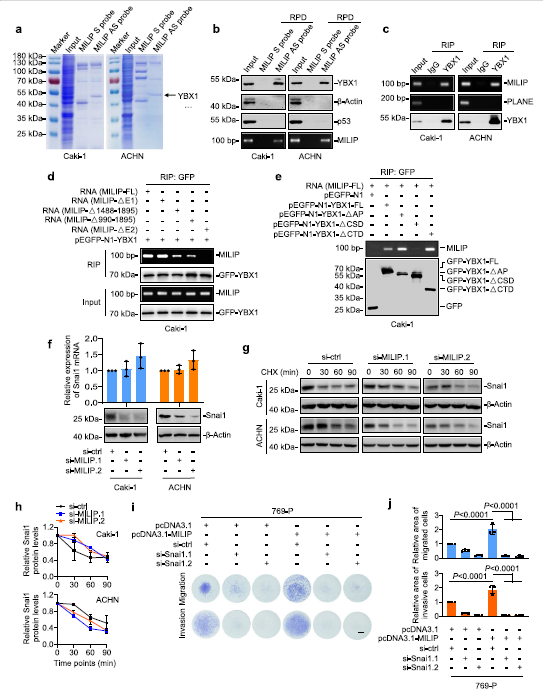
3)MILIP與YBX1相互作用,促進(jìn)Snai1的翻譯激活
為了了解ccRCC細(xì)胞中MILIP介導(dǎo)的EMT促進(jìn)機(jī)制,我們使用RNA下拉和質(zhì)譜分析確定了與MILIP相互作用的蛋白質(zhì)。與MILIP共沉淀的最豐富的蛋白質(zhì)是YBX1(圖3a),這是一種核酸結(jié)合蛋白,已知可翻譯激活Snai1,通過轉(zhuǎn)錄抑制下調(diào)E-cadherin表達(dá),從而促進(jìn)EMT。在Caki-1和ACHN細(xì)胞中使用RNA下拉和RIP測(cè)定確認(rèn)了MILIP和YBX1之間的關(guān)聯(lián)(圖3b,c)。為了確定負(fù)責(zé)與YBX1相互作用的MILIP區(qū)域,我們對(duì)體外轉(zhuǎn)錄的MILIP突變體進(jìn)行了映射實(shí)驗(yàn)。該分析表明,位于外顯子2 (E2)內(nèi)的MILIP片段618-989是其與YBX1結(jié)合的關(guān)鍵(圖3d)。另一方面,YBX1的CSD缺失減少了其與MILIP的關(guān)聯(lián)(圖3e)。值得注意的是,MILIP的敲低導(dǎo)致Snai1蛋白水平的下調(diào),而不影響Snai1 mRNA表達(dá)(圖3f)。環(huán)己酰亞胺追蹤實(shí)驗(yàn)表明,MILIP的敲除不會(huì)改變Snai1蛋白的周轉(zhuǎn)率(圖3g,h)。Snai1的敲低消除了MILIP過表達(dá)誘導(dǎo)的ccRCC細(xì)胞遷移和侵襲(圖3i,j),表明Snai1對(duì)于MILIP介導(dǎo)的ccRCC細(xì)胞轉(zhuǎn)移的促進(jìn)至關(guān)重要。
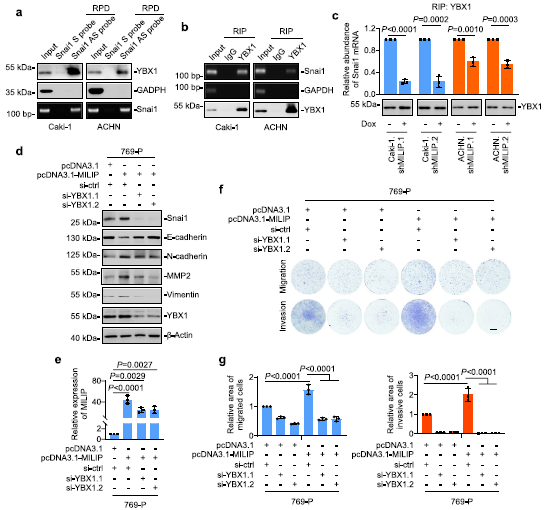
4)MILIP與Snai1 mRNA形成RNA-RNA雙鏈
我們研究了MILIP如何促進(jìn)Snai1的翻譯激活。使用IntaRNA程序進(jìn)行的生物信息學(xué)分析確定了Snai1 mRNA的5′UTR附近的潛在MILIP結(jié)合區(qū)(MILIP-BR),該區(qū)域與MILIP中富含 DFO的片段互補(bǔ)。體外合成的生物素標(biāo)記的MILIP RNA沉淀了含有MILIP-BR的RNA片段(圖4a)。然而,當(dāng)MILIP內(nèi)富含DFO的片段被刪除時(shí),這種關(guān)聯(lián)性減弱(圖4a)。因此,MILIP和Snai1 mRNA分別通過DFO和MILIP-BR形成RNA-RNA雙鏈體。作為支持,內(nèi)源性MILIP從Caki-1和ACHN細(xì)胞提取物中沉淀Snai1 mRNA(圖4b)。我們還測(cè)試了RNA-RNA雙鏈在MILIP介導(dǎo)的Snai1翻譯調(diào)控中的功能重要性。與野生型MILIP的過表達(dá)(圖4c)相反,將缺乏DFO的MILIP突變體(MILIP-ΔDFO)轉(zhuǎn)染769-P細(xì)胞不會(huì)影響Snai1蛋白表達(dá)水平,也不會(huì)影響細(xì)胞遷移和侵襲(圖4c-e)。因此,RNA-RNA雙鏈的形成是MILIP介導(dǎo)的Snai1翻譯促進(jìn)所必需的。

5)MILIP將YBX1與Snai1的翻譯激活聯(lián)系起來
我們接下來研究了MILIP和YBX1在調(diào)節(jié)Snai1 mRNA翻譯中的關(guān)系。正如預(yù)期的那樣,YBX1與Snai1 mRNA結(jié)合(圖5a,b)。然而,在MILIP敲低的細(xì)胞中,YBX1和Snai1 mRNA之間的這種關(guān)聯(lián)減少(圖5c),表明MILIP促進(jìn)YBX1與Snai1 mRNA的關(guān)聯(lián)。一致地,在769-P細(xì)胞中,YBX1的敲除消除了MILIP過表達(dá)觸發(fā)的Snai1、N-cadherin、MMP2和波形蛋白的上調(diào)和E-cadherin的下調(diào)(圖5d,e)。此外,YBX1的敲除至少部分阻斷了769-P細(xì)胞中MILIP介導(dǎo)的遷移和侵襲增加(圖5f,g)。總之,這些結(jié)果表明MILIP促進(jìn)YBX1與Snai1 mRNA的結(jié)合,并促進(jìn)YBX1介導(dǎo)的Snai1的翻譯激活。
結(jié)論:
TFAP2C響應(yīng)的MILIP通過與Snai1 mRNA形成RNA-RNA雙鏈以促進(jìn)YBX1介導(dǎo)的Snai1翻譯激活,促進(jìn)ccRCC轉(zhuǎn)移。MILIP在轉(zhuǎn)移性ccRCCs中上調(diào),且MILIP高表達(dá)與ccRCC患者預(yù)后不良相關(guān)。這些發(fā)現(xiàn)揭示了lncRNA介導(dǎo)的促進(jìn)ccRCC轉(zhuǎn)移的機(jī)制,并暗示靶向MILIP代表了治療晚期ccRCC的新途徑。
參考文獻(xiàn):
Wang Y, Feng YC, Gan Y, Teng L, Wang L, La T, Wang P, Gu Y, Yan L, Li N, Zhang L, Wang L, Thorne RF, Zhang XD, Cao H, Shao FM. LncRNA MILIP links YBX1 to translational activation of Snai1 and promotes metastasis in clear cell renal cell carcinoma. J Exp Clin Cancer Res. 2022 Aug 26;41(1):260. doi: 10.1186/s13046-022-02452-9.