






CircMYH9通過調節絲氨酸代謝和氧化還原穩態來驅動結直腸癌生長
環狀RNA(circRNA)在癌癥進展和代謝調節中發揮重要作用。絲氨酸/甘氨酸代謝通過促進癌細胞的合成代謝需求和表觀基因組以及調節其氧化還原狀態來支持癌細胞的生長。然而,circRNA在調節絲氨酸/甘氨酸(SG)代謝中的作用尚未得到很好的闡明。今天講一篇關于circRNA調節絲氨酸/甘氨酸代謝的文章,改文章發表于Molecular Cancer期刊,影響因子27.4。文章題名為:CircMYH9 drives colorectal cancer growth by regulating serine metabolism and redox homeostasis in a p53-dependent manner。
在結直腸癌組織和細胞中鑒定出circMYH9
為了識別潛在的差異表達的 circRNA,從 GEO 數據庫中篩選了可能在 CRC和癌旁組織中差異表達的circRNA。共檢測到8個下調的 circRNA和237 個上調的circRNA。然后,通過微陣列分析進一步分析3對CRC和癌旁組織中的circRNA表達譜,92個circRNA上調,85個circRNA下調。數據庫中的 237 個上調circRNA與微陣列中92個上調circRNA相交,鑒定出9個重疊的 circRNA。在9個circRNA中,hsa_circ_0092283,也稱為 circMYH9,起源于 MYH9 基因,由內含子37的頭尾剪接組成。由于circMYH9與其他8個circRNA相比顯著上調,選擇它進行進一步研究。qRT-PCR 分析表明 circMYH9可以通過cDNA中的不同引物進行擴增,并且在基因組 DNA(gDNA)組中沒有觀察到其他產物。此外,circMYH9 對 RNase R具有抗性,而 MYH9 mRNA 可以被RNase R降解。然后,通過 Sanger 測序證實了circMYH9 的RT-PCR 產物。

CircMYH9 表達在 CRC 組織中上調并預測預后不良
qRT-PCR和FISH檢測CRC組織中的circMYH9表達水平,顯示CRC組織中的circMYH9表達高于腫瘤周圍組織。然后,檢查了148例CRC病例中 circMYH9表達與臨床病理結果的相關性。circMYH9 水平與腫瘤大小、遠處轉移、淋巴結轉移、TNM 分期和 p53 狀態顯著相關。circMYH9低表達患者的無復發生存率(RFS)高于高表達患者,風險比為 0.593。此外,circMYH9高表達患者的總生存率(OS)顯著低于circMYH9低表達患者,風險比為 0.547。
CircMYH9促進CRC細胞中的細胞生長
首先檢查了其在正常腸上皮細胞系FHC和CRC細胞系中的表達,發現與 FHC細胞相比,circMYH9在CRC 細胞系中的表達顯著上調。CCK-8和集落形成測定表明,circMYH9敲低導致HCT116和HCT8細胞中腫瘤生長的顯著抑制。circMYH9過表達增加了LoVo細胞的生長。有趣的是,與上述 p53 野生型 (wt) CRC細胞系相比,circMYH9 敲低僅略微改變了p53突變的DLD1和HT-29細胞系中的細胞增殖。細胞周期分布分析表明,circMYH9敲低導致細胞周期在G0/G1期停滯,S期和G2/M期細胞百分比相應降低。circMYH9敲低后,細胞周期蛋白(CDK1、Cyclin D1、CDK6、Cyclin E2)顯著減少,而在CRC細胞中circMYH9過表達后,細胞周期得到促進,細胞周期蛋白增加。由于 HCT116 和 LoVo 細胞系在 p53 wt CRC 細胞系中分別相對較高和較低,我們選擇它們進行進一步研究。

接下來檢查了circMYH9對體內CRC腫瘤生長的影響。將circMYH9 shRNA或shCtrl對照穩定轉染的HCT116細胞和circMYH9質粒或載體對照穩定轉染的LoVo細胞皮下注射到裸鼠體內,注射21天后測量腫瘤體積,處死后測量腫瘤重量。與 shCtrl 對照細胞相比,CircMYH9敲除顯著抑制了腫瘤體積和重量,相比之下,與載體對照細胞相比,CircMYH9 過表達表現出增加的腫瘤體積和重量。IHC 分析顯示,由 circMYH9 shRNA 轉染的 HCT116 細胞形成的腫瘤表現出較弱的 Ki67 染色和由過度表達 circMYH9 的 LoVo 細胞形成的細胞表現出比源自載體轉染細胞的腫瘤更強的 Ki67 染色。
CircMYH9通過p53介導的PHGDH上調促進SG代謝
為了深入了解 circMYH9 促進結直腸腫瘤發生的分子機制,在對照和敲除 circMYH9的HCT116細胞中進行了RNA-seq,并鑒定了差異表達的基因。CircMYH9 缺失導致 893 個基因的上調和 1650 個基因的下調。然后,使用基因本體(GO)分析和GSEA分析進行功能富集分析。前20個GO術語顯示包括甘氨酸、絲氨酸和蘇氨酸代謝和p53信號通路。GSEA結果顯示差異表達的基因集與SG代謝和p53通路顯著相關。沉默或過表達circMYH9,發現它正調節SG生物合成途徑基因(PHGDH、PSAT1、PSPH和SHMT2)的表達。circMYH9 的沉默或過表達幾乎不會改變中性氨基酸轉運蛋白 SLC1A4 的水平,表明 circMYH9 不能影響SG的攝取。在培養的最后一個小時,將表達對照或 circMYH9-shRNA 的 HCT116 細胞和表達載體或 circMYH9 質粒的 LoVo 細胞與均勻標記的 [U-13C] 葡萄糖一起溫育,并通過液相色譜-質譜法檢測。結果顯示,與對照細胞相比,shRNA 轉染的 HCT116 細胞中的SG顯著降低,而過表達circMYH9的 LoVo 細胞中的 SG 相對于載體細胞顯著增加。
為了驗證circMYH9 是否通過 p53 途徑調節SG的合成。qRT-PCR和WB 證明了circMYH9 對p53的負調節。Nutlin-3可激活 p53。Nutlin-3處理過表達 circMYH9的 LoVo 細胞逆轉了p53和MDM2的下調,并抑制了SG生物合成途徑基因表達和細胞增殖。而circMYH9耗盡的HCT116細胞中的p53沉默則顯示出相反的效果。還在過度表達 circMYH9 的 LoVo 細胞中用 shRNA 敲低了 PHGDH。PHGDH 抑制顯示出與 Nutlin-3 處理類似的效果,但不影響 p53 表達。此外,與對照細胞相比,具有 circMYH9 敲除的體內異種移植物的 IHC 顯示出更高的 p53 表達和更低的 PHGDH 和 PSPH 表達。相比之下,circMYH9 過表達異種移植物的 IHC 顯示 p53 的表達較低,而 PHGDH 和 PSPH 的表達高于載體細胞的表達。這些結果證實了 circMYH9 在 p53 通路和從頭 SG 代謝的調節中的主要作用。

CircMYH9 通過 hnRNPA2B1 調節 p53 前體 mRNA 的穩定性
為了解釋 circMYH9 調節p53表達的機制,進行了qRT-PCR和FISH以確定circMYH9在HCT116和LoVo 細胞中的亞細胞位置。結果表明,circMYH9主要定位于細胞核中。由于circMYH9敲低不影響p53啟動子活性。有趣的是,在用放線菌素D阻斷新的RNA合成后,發現沉默 circMYH9 并沒有在 12 小時內改變p53 mRNA 的穩定性,但在24小時內顯著增強了p53 mRNA的穩定性,表明 circMYH9可能間接抑制p53的mRNA降解。事實上,circMYH9 siRNA處理后p53前體mRNA表達增加,circMYH9質粒處理后p53前體mRNA表達降低。結果表明,circMYH9 的缺失可能導致 p53 前體 mRNA 的增加,進而影響 p53 的 mRNA 表達。
通過circMYH9驗證了某些蛋白質是否參與調節p53前mRNA的穩定性。ChIP結合MS用于鑒定circMYH9的結合蛋白,在此過程中鑒定了42種蛋白質,包括 hnRNPA2B1。為了驗證MS結果,進行了RIP測定并觀察到circMYH9在由hnRNPA2B1抗體沉淀的復合物中的富集。hnRNPA2B1和circMYH9之間的相互作用被非生物素化的 circMYH9 以劑量依賴性方式競爭性抑制。免疫熒光共定位分析顯示 circMYH9 和 hnRNPB2A1 共定位在 CRC 細胞的細胞核中、皮爾遜相關分析(Rx)和相關系數分析(R)顯示顯示高度相關。
為了檢查hnRNPA2B1和p53前體mRNA的關聯,進行使用抗 hnRNPA2B1的 RIP測定,證明 hnRNPA2B1與p53前體mRNA相互作用。使用生物素化的反義 DNA 探針提取了p53前體mRNA。hnRNPA2B1被共沉淀。hnRNPA2B1 敲低導致 p53 的前體 mRNA 和蛋白質水平降低。這些結果表明 hnRNPA2B1 可能對p53前體 mRNA 發揮穩定作用。qPCR結果顯示hnRNPA2B1敲低可以消circMYH9缺失對p53 pre-mRNA表達的促進作用,而 hnRNPA2B1 過表達可以挽救由circMYH9過表達誘導的p53前體 mRNA的下調表達。circMYH9 敲低增加了hnRNPA2B1和p53前體mRNA之間的結合,而增加的 circMYH9 表達減弱了 hnRNPA2B1 與p53前體mRNA的結合。這些發現表明circMYH9通過阻止hnRNPA2B1與p53前體 mRNA 結合來抑制p53前體mRNA 的穩定性。

p53 3' 非翻譯區 (UTR) 介導的 hnRNPA2B1 對前 mRNA 穩定性的調節
為了進一步確定 p53 的哪個區域參與穩定性調節,在 HCT116 細胞中進行了 MeRIP-seq,發現一個顯著的m6A峰分布在 p53 mRNA 的 3'UTR。構建了一個帶有 Flag 標簽的表達載體,包括帶有 3'UTR (p53 CDS-3'UTR) 或單獨 CDS (p53 CDS)的p53編碼序列,以及用 hnRNPA2B1 siRNA 共轉染的 HCT116 細胞。結果表明,hnRNPA2B1 敲低顯著降低了p53 CDS-3'UTR轉染的CRC細胞中 p53 的表達,而不是單獨的 p53 CDS。這表明 3'UTR 對于 hnRNPA2B1 介導的p53調節是必不可少的。
結果顯示,敲低Mettl3降低了HCT116細胞中p53前體mRNA水平,而 Mettl3 過表達增加了p53前體mRNA水平而不影響hnRNPA2B1的蛋白質水平。使用針對 p53 3'UTR 和CDS區域的特異性引物進行MeRIP-qPCR以檢測 m6A水平,結果顯示沉默 Mettl3 導致3' UTR峰m6A甲基化水平降低,而過表達 Mettl3 具有相反的效果。接下來,RIP分析證實了hnRNPA2B1和p53前體 mRNA 之間的相互作用通過消耗Mettl3被抑制或通過過度表達Mettl3 增強。
p53 3'UTR 熒光素酶報告基因檢測發現hnRNPA2B1敲低降低了含有 p53 3'UTR的熒光素酶構建體的活性,而Mettl3過表達增加了由 hnRNPA2B1 敲低誘導的熒光素酶活性降低。建立了 p53 3'UTR mut 熒光素酶測定。發現p53 3'UTR mut的螢光素酶活性對hnRNPA2B1敲低或Mettl3過表達沒有反應。數據表明,p53前體mRNA的表達受hnRNPA2B1與p53 3'UTR上m6A位點的結合控制。

CircMYH9在氨基酸剝奪下被ROS-HIF1α通路激活
癌細胞可能會暫時或永久地進入非必需的營養缺陷狀態,檢測了氨基酸缺乏是否與CR細胞中的circMYH9過表達有關。我們在缺乏SG或缺乏谷氨酰胺的培養基中培養CRC細胞,并且 circMYH9 響應于SG或Glu剝奪而上調。缺乏 Glu 或 SG 會導致 ROS 水平升高,從而導致 HIF1α增加。使用ROS清除劑 NAC來處理缺乏SG或Glu的細胞。正如預期的那樣,NAC在不含SG或不含 Glu的培養基中逆轉了HIF1α和circMYH9的升高。HIF1α敲除消除了HCT116 和LoVo細胞中Glu或SG剝奪誘導的circMYH9表達。表明SG或Glu剝奪誘導的細胞ROS積累增加了HIF1α水平,從而促進了circMYH9的表達。
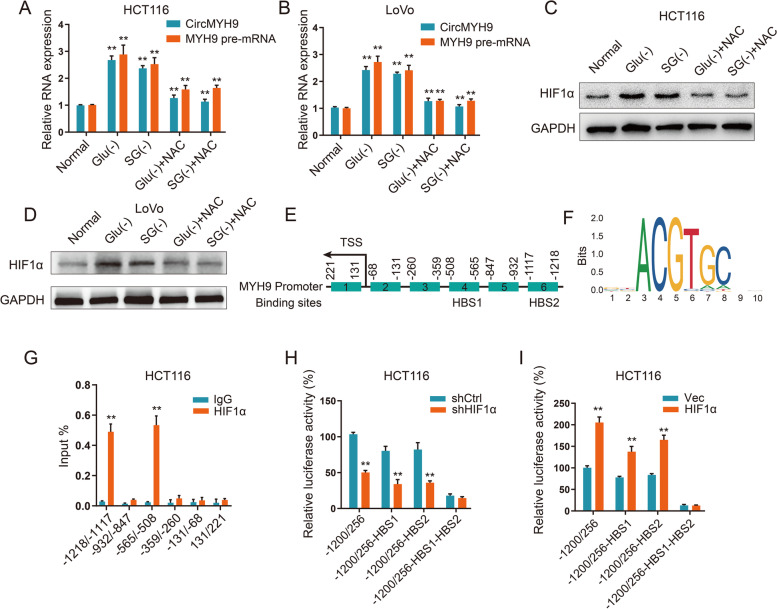
為了探索 HIF1α促進circMYH9表達的分子機制,測量了 MYH9 前體 mRNA 的表達。SG或Glu饑餓增加了MYH9的前體mRNA 水平,HIF1α敲低降低了缺乏SG或Glu的CRC細胞中的MYH9前體 mRNA 水平。這些結果表明HIF1α在轉錄水平上促進 MYH9 前體 mRNA,這進一步增加了circMYH9。使用 JASPAR 搜索了MYH9啟動子中的HIF1α結合位點和基序。確定了兩個最可能的HIF1α結合位點。ChIP-qPCR 證明 HIF1α與HCT116細胞中MYH9 啟動子的兩個HBS位點結合。為了證實HIF1α通過HBS促進MYH9表達,構建了包含全長MYH9啟動子區域和HBS缺失的circMYH9啟動子區域的熒光素酶報告基因。HBS1或HBS2的缺失部分消除了HIF1α敲低或過表達對MYH9 熒光素酶報告基因活性的抑制或促進,而HBS1和HBS2的缺失則完全消除了 HIF1α shRNA 或質粒對MYH9熒光素酶報告基因活性的影響。這些數據表明,HBS區域對于SG或Glu剝奪下的HIF1α依賴性circMYH9轉錄調控至關重要。
CircMYH9 促進 p53 野生型小鼠的結直腸腫瘤發生
為了確定 circMYH9 在體內 CRC 發病機制中的因果作用,通過將 p53 fl/fl小鼠與Villin-Cre小鼠雜交,產生了結腸特異性、條件性 p53 KO小鼠。p53 WT小鼠和 p53 KO小鼠用氧化偶氮甲烷 (AOM) 和葡聚糖硫酸鈉 (DSS) 處理59 天以誘導結直腸腫瘤發生。在AOM注射前一周,使用AAV9作為載體在小鼠結腸中特異性過表達circMYH9。AAV-circMYH9 的轉染效率通過qPCR驗證。與p53WT小鼠相比,p53KO小鼠的腫瘤數量、發病率和大小更多,與AAV-ctrl治療的p53WT小鼠相比,p53WT小鼠中circMYH9的過度表達導致腫瘤數量、發病率和大小增加。帶有針對 circMYH9 的探針 FISH證實circMYH9仍然在p53 wt + AAV 小鼠的腫瘤組織中表達。IHC 檢查顯示,與 ctrl-AAV小鼠相比,具有 circMYH9過表達的腫瘤顯示出p53的表達受到抑制,而PHGDH和PSPH的表達增加。總之,這些結果表明 circMYH9 可以通過抑制 p53 和促進小鼠 SG 代謝來驅動化學誘導的致癌作用。

該研究首次證明內含子衍生的circMYH9通過調節SG代謝和氧化還原穩態促進CRC的生長。還確定p53作為circMYH9的靶標,并表明circMYH9可以通過募集m6A閱讀器hnRNPA2B1降解 p53前體 mRNA來抑制 p53,進一步轉錄抑制 PHGDH 表達。此外,在 SG或Glu饑餓下鑒定了擴增circMYH9的 ROS/HIF1α途徑。工作突出了CRC中circMYH9的新機制,為監測和治療CRC 提供了寶貴的資源。