






lncRNA MIR210HG促進子宮內(nèi)膜癌的進展
上皮-間充質(zhì)轉(zhuǎn)化(EMT)促進腫瘤發(fā)生和轉(zhuǎn)移,增加腫瘤對治療干預(yù)的耐受性。TGF-b和Wnt通路的異常激活誘導(dǎo)EMT。lncRNAs顯著影響EMT調(diào)控。2021年6月發(fā)表于Molecular Therapy-Nucleic Acids(IF=8.886)的文章“lncRNA MIR210HG promotes the progression of endometrial cancer by spongingmiR-337-3p/137 via the HMGA2-TGF-b/Wnt pathway”對此展開了研究。在此,我們發(fā)現(xiàn)MIR210HG在子宮內(nèi)膜癌組織中過表達,與不良預(yù)后相關(guān)。MIR210HG的沉默在體外顯著抑制了細胞增殖、遷移、侵襲和EMT表型的形成以及在體內(nèi)的腫瘤發(fā)生。生物信息學(xué)分析、RIP分析和熒光素酶分析表明,MIR210HG作為miR-337-3p和miR-137的分子海綿,調(diào)節(jié)HMGA2的表達。此外,MIR210HG過表達顯著增強了Wnt/b-catenin和TGF-b/Smad3信號通路基因,而MIR210HG或HMGA2下調(diào)抑制了Wnt/b-catenin和TGF-b/Smad3信號通路基因。我們在MIR210HG-miR-337-3p/137-HMGA2軸上的發(fā)現(xiàn)說明了其作為子宮內(nèi)膜癌治療發(fā)展的靶點的潛力。

技術(shù)路線

結(jié)果
1)MIR210HG與子宮內(nèi)膜癌患者的低生存率相關(guān)
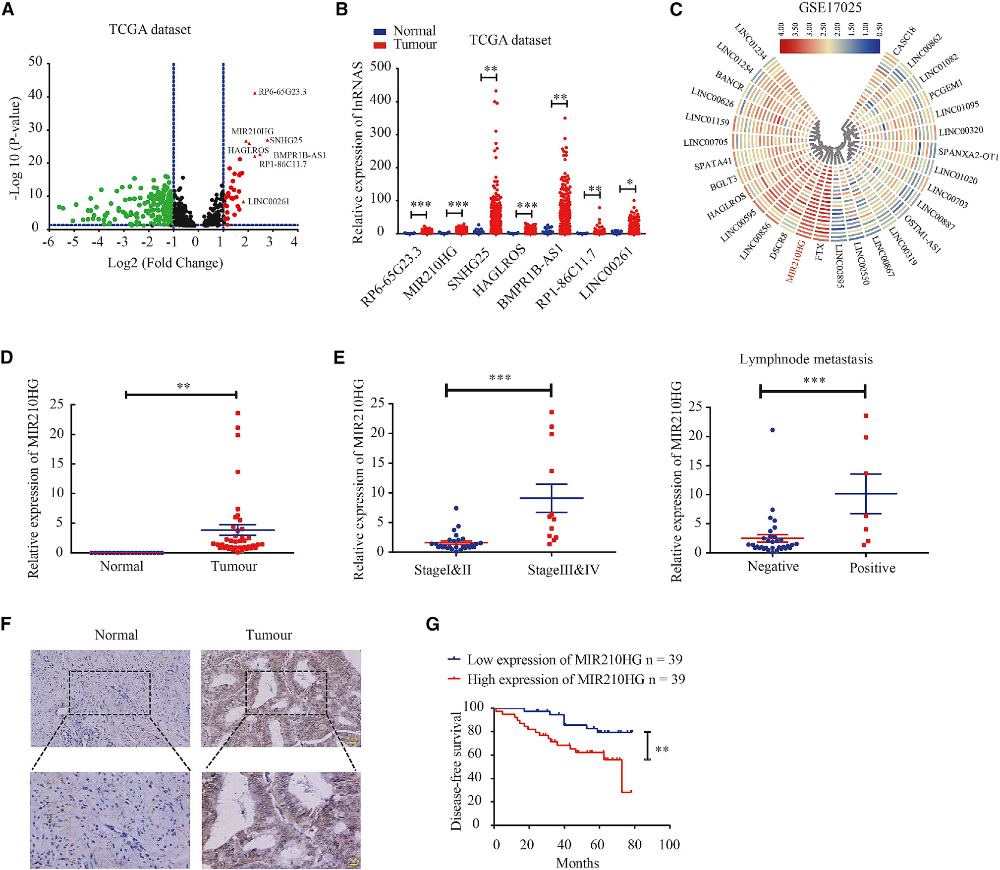
為了識別子宮內(nèi)膜癌組中異常表達的lncRNA,我們分析了來自TCGA的數(shù)據(jù)。表達陣列分析顯示332個lncRNA的表達差異有統(tǒng)計學(xué)意義。7個lncRNA表達上調(diào)(圖1A和1B)。然后,我們分析了GEO數(shù)據(jù)集,發(fā)現(xiàn)在子宮內(nèi)膜癌GSE17025隊列中MIR210HG的表達也上調(diào)(圖1C)。qPCR顯示MIR210HG在子宮內(nèi)膜癌中的表達明顯高于正常子宮內(nèi)膜組織(圖1D)。此外,我們分析了MIR210HG的表達與子宮內(nèi)膜癌患者臨床病理參數(shù)的關(guān)系,發(fā)現(xiàn)MIR210HG在從I、II期到III、IV期的進展過程中增加。MIR210HG的表達與淋巴結(jié)轉(zhuǎn)移顯著相關(guān)(圖1E)。原位雜交實驗結(jié)果顯示,MIR210HG在子宮內(nèi)膜癌中的表達水平高于正常子宮內(nèi)膜組織(圖1F)。MIR210HG高表達的子宮內(nèi)膜癌患者生存率較低(圖1G)。
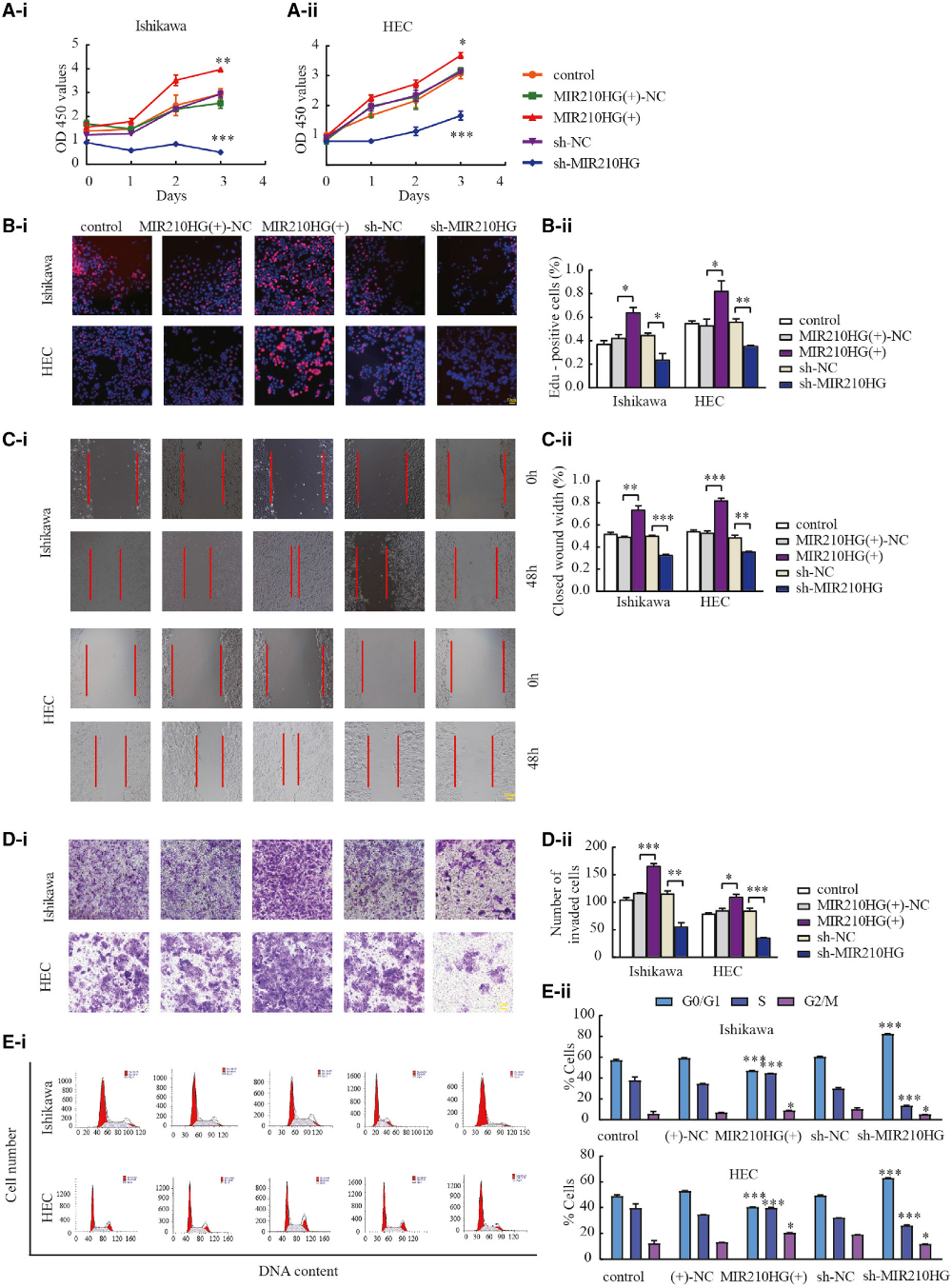
2)下調(diào)MIR210HG可抑制子宮內(nèi)膜癌細胞的增殖、遷移和侵襲
我們研究了MIR210HG在子宮內(nèi)膜癌細胞中的生物學(xué)功能。我們發(fā)現(xiàn)MIR210HG的下調(diào)有效地降低了Ishikawa和HEC-1A細胞的增殖(圖2A和2B)。采用創(chuàng)面愈合實驗和Transwell實驗研究MIR210HG對腫瘤遷移和侵襲的影響。與sh陰性對照組(NC)相比,轉(zhuǎn)染sh-MIR210HG的細胞遷移和侵襲減少(圖2C和2D)。流式細胞術(shù)分析細胞周期分布(圖2E)。以上結(jié)果提示MIR210HG在子宮內(nèi)膜癌中起致癌基因的作用。
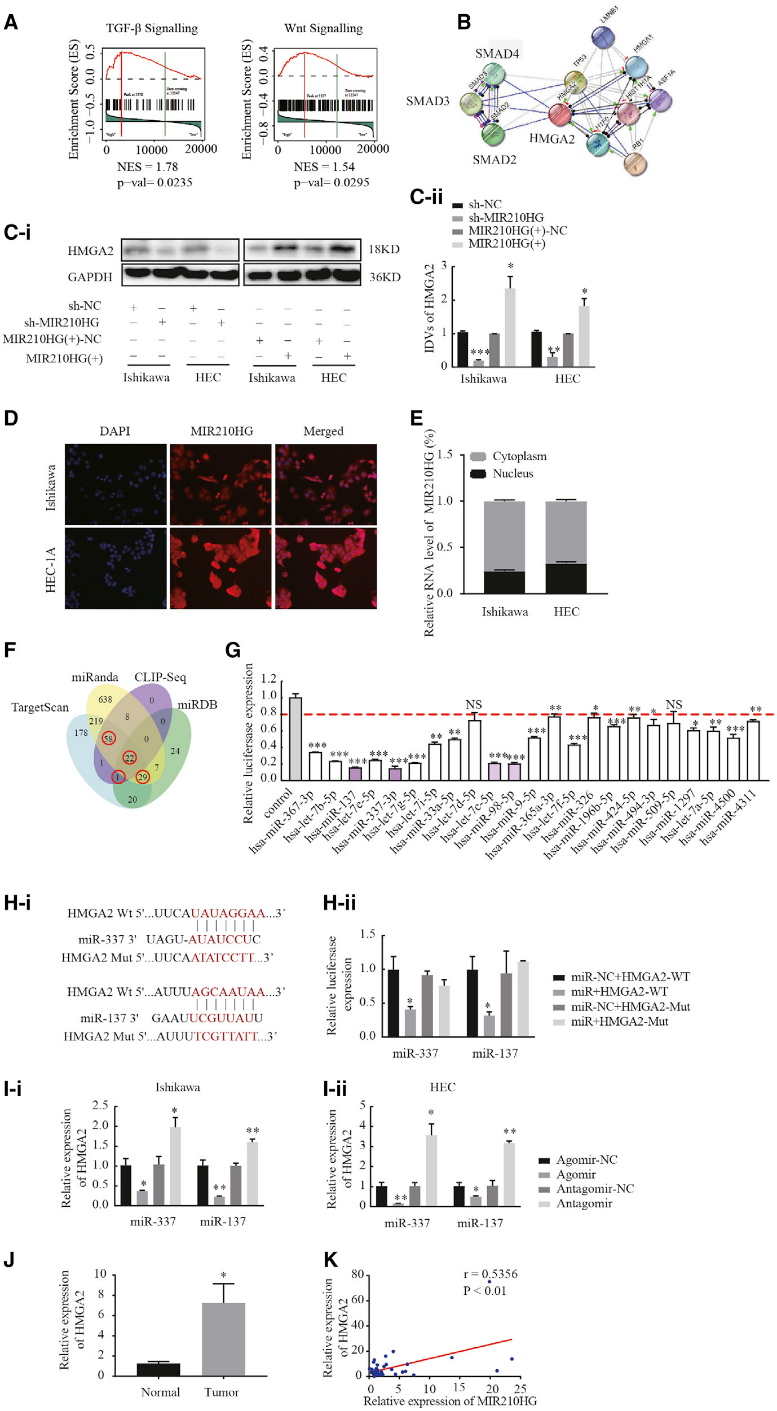
3)MIR210HG可能參與了由lncRNA-miRNA-mRNA調(diào)控網(wǎng)絡(luò)介導(dǎo)的子宮內(nèi)膜惡性生物學(xué)行為
利用TCGA數(shù)據(jù)集對子宮內(nèi)膜癌樣本進行基因集合富集分析,探索MIR210HG參與調(diào)控子宮內(nèi)膜癌的生物學(xué)通路。MIR210HG的表達與TGF-b和Wnt通路顯著相關(guān),而Smad3基因在這些關(guān)聯(lián)中發(fā)揮了重要作用(圖3A)。搜索工具STRING的分析也顯示SMAD3和HMGA2是強相關(guān)的(圖3B)。我們探究HMGA2是否為MIR210HG的下游基因,發(fā)現(xiàn)MIR210HG下調(diào)后HMGA2蛋白表達水平降低。相反,當MIR210HG高表達時,HMGA2的表達顯著增加(圖3C)。這表明MIR210HG可能通過上調(diào)HMGA2的表達來促進子宮內(nèi)膜癌細胞的惡性生物學(xué)行為。熒光原位雜交和核胞質(zhì)RNA分數(shù)實驗結(jié)果顯示,MIR210HG在Ishikawa細胞和HEC-1A細胞中均位于核和細胞質(zhì)內(nèi)(圖3D和3E)。我們假設(shè)MIR210HG對miRNAs起海綿作用。為了探究MIR210HG是否參與了一個新的MIR210HG- miRNA -HMGA2調(diào)控網(wǎng)絡(luò),我們使用TargetScan、miRanda、CLIP-Seq和miRDB生物信息學(xué)數(shù)據(jù)庫來預(yù)測與HMGA2結(jié)合的mirna(圖3F)。選擇3個軟件程序分析交叉處的miRNAs作為候選miRNAs。通過將野生型雙熒光素酶載體介導(dǎo)的HMGA2構(gòu)建物共轉(zhuǎn)染HEK293T細胞,共鑒定和篩選了110個miRNAs。轉(zhuǎn)染后,23個miRNA轉(zhuǎn)染細胞的熒光素酶活性顯著降低(圖3G)。在上述23種miRNAs中,轉(zhuǎn)染miR-3373p、miR-137、let-7c-5p和miR-98-5p時,熒光素酶活性顯著降低,其中轉(zhuǎn)染miR-337-3p和miR-137時,熒光素酶活性降低最多。利用生物信息學(xué)數(shù)據(jù)庫TargetScan預(yù)測miR-337-3p和miR-137中與HMGA2結(jié)合的位點。雙熒光素酶基因報告基因檢測結(jié)果顯示,miR-337-3p和miR-137在其預(yù)測的結(jié)合位點上結(jié)合HMGA2(圖3H)。qPCR結(jié)果顯示,miR-337-3p和miR-137負調(diào)控HMGA2的表達(圖3I)。采用Pearson’s秩相關(guān)法分析MIR210HG與HMGA2表達的相關(guān)性,發(fā)現(xiàn)HMGA2的表達與MIR210HG呈正相關(guān)(圖3 J和K)。這些結(jié)果表明MIR210HG可能參與了新的MIR210HG-miR-337-3p/137-HMGA2調(diào)控網(wǎng)絡(luò)。
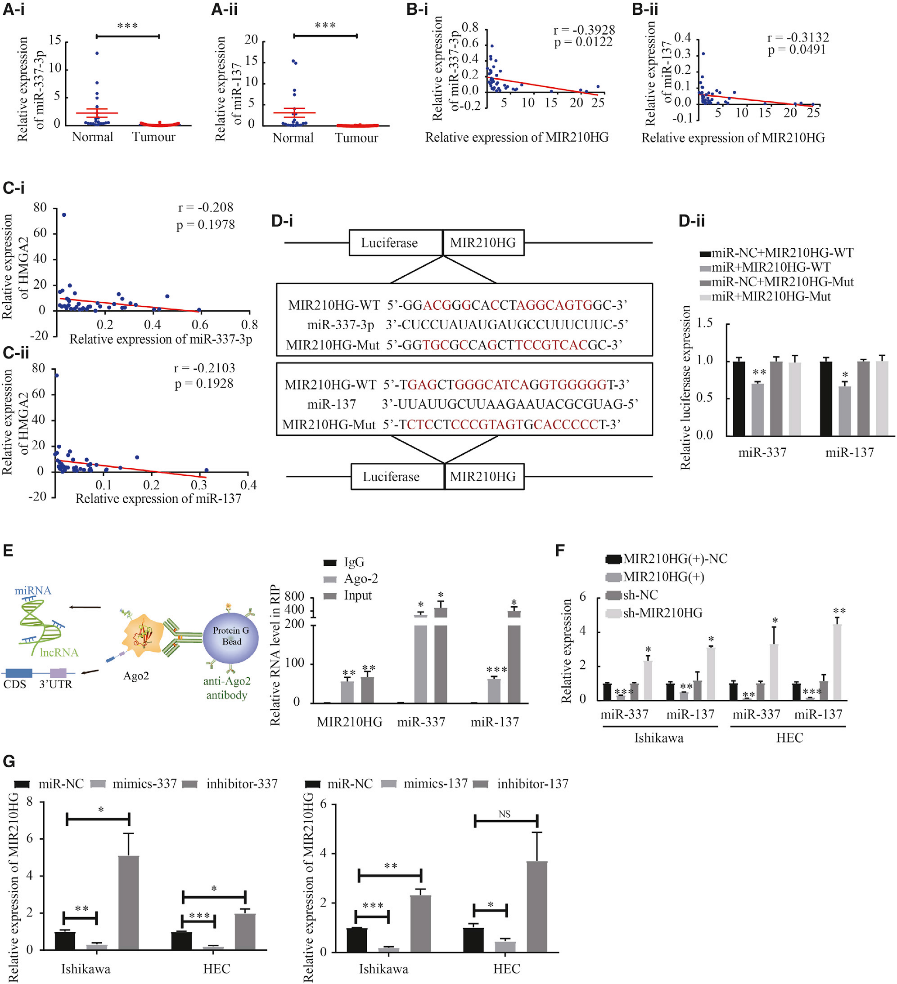
4)miR-337-3p/miR-137在子宮內(nèi)膜癌組織中表達較低,miR-337-3p/miR-137是MIR210HG的靶點
qPCR結(jié)果顯示,腫瘤組織中miR-337-3p和miR-137的表達明顯下調(diào)(圖4A)。此外,采用Pearson等級相關(guān)法分析miR-337-3p、miR-137與MIR210HG表達的相關(guān)性(圖4B)。隨后,我們分析了HMGA2與miR-337-3p/137表達的關(guān)系(圖4C)。我們進行了熒光素酶實驗,以確定MIR210HG是否可以直接靶向miR-337-3p和miR-137在預(yù)測的結(jié)合位點(圖4D)。為了確定MIR210HG是否與miRNA核糖核蛋白復(fù)合物結(jié)合,我們使用抗AGO2抗體進行RIP實驗。與對照免疫球蛋白G 免疫沉淀相比,lncRNA MIR210HG和miR-337-3p/miR-137在含AGO2的免疫沉淀中顯著富集(圖4E)。當MIR210HG在Ishikawa和HEC-1A細胞中過表達時,miR-337-3p和miR-137表達水平降低。相比之下,轉(zhuǎn)染sh-MIR210HG后,細胞中miR-337-3p和miR-137的表達水平顯著升高,說明MIR210HG可以調(diào)控miR-337-3p和miR-137的表達(圖4F)。隨后,我們研究了miR-337-3p和miR-137是否負調(diào)控MIR210HG的表達,發(fā)現(xiàn)過表達miR-337-3p/137會下調(diào)MIR210HG的表達,而敲除miR-337-3p/137會上調(diào)MIR210HG的表達(圖4G)。
5)過表達miR-337-3p和miR-137抑制子宮內(nèi)膜癌細胞的惡性生物學(xué)行為
為了研究miR-337-3p和miR-137在子宮內(nèi)膜癌生物學(xué)行為中的可能作用,我們分別用agomir-337-3p/137和antagomir-3373p/137轉(zhuǎn)染Ishikawa和HEC-1A細胞。使用CCK-8和EdU檢測細胞增殖(圖5A和5B)。傷口愈合和Transwell檢測分別用于評估細胞遷移和侵襲。過表達miR-33373p和miR-137降低了Ishikawa和HEC-1A細胞的遷移和侵襲能力(圖5C和5D)。流式細胞術(shù)檢測并分析細胞周期分布(圖5E)。以上實驗結(jié)果證實,與miR-NC組相比,miR-337-3p和miR-137過表達顯著抑制了細胞的增殖、侵襲和遷移。
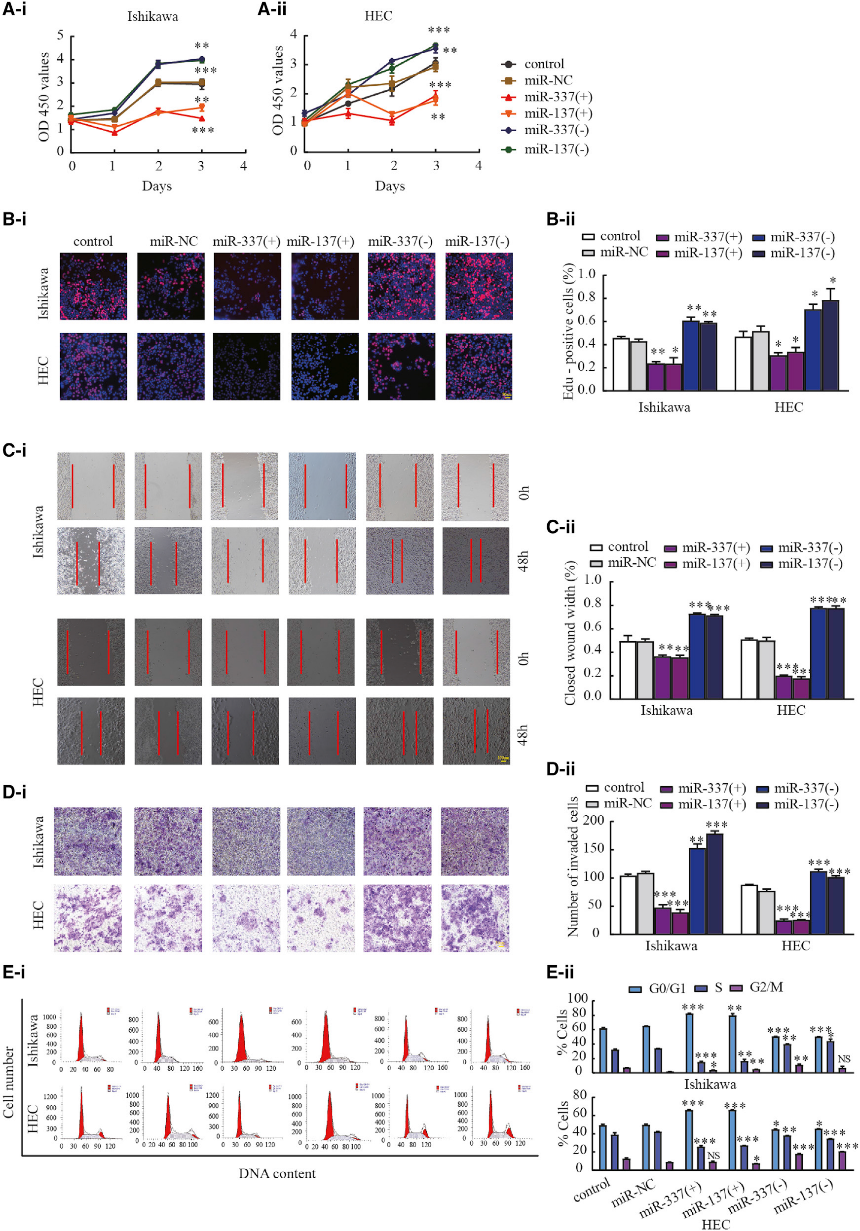
6)沉默MIR210HG可以通過miR-337-3p/137-HMGA2軸阻斷TGF-b和Wnt信號通路
之前的GSEA表明MIR210HG與TGFb/Smad3和Wnt/b-catenin信號通路相關(guān)。為了進一步研究MIR210HG調(diào)控子宮內(nèi)膜癌細胞進展的機制,我們檢測了MIR210HG、HMGA2、miR-337-3p和miR-137對TGF-b、Wnt和EMT通路相關(guān)蛋白水平的調(diào)控作用。MIR210HG的下調(diào)降低了TGF-bII、p-Smad3和Snail的表達以及b-catenin在細胞核中的分布(圖6A和6B)。基于這些結(jié)果,我們推測MIR210HG可能通過靶向miR-3373p/137-HMGA2調(diào)控軸抑制EMT、TGF-b和Wnt信號通路。干擾HMGA2顯著降低TGF-bII、p-Smad3和Snail的表達,b-catenin在細胞核中的分布(圖6C和6D)。我們之前曾報道過HMGA2在Ishikawa和HEC-1A細胞系中調(diào)節(jié)Snail、Slug、N-cadherin、MMP-2和MMP-9的表達。我們接下來研究了miR-337-3p/137表達的誘導(dǎo)是否影響TGF-b、Wnt和EMT通路相關(guān)蛋白的水平。結(jié)果顯示,過表達miR-337-3p和miR-137抑制了HMGA2、TGF-bII、p-Smad3和Snail的表達(圖6E和6F)。agomir-3373p/137處理和sh-MIR210HG-或sh- HMGA2處理Ishikawa和HEC-1A細胞中,b-catenin在細胞核中的分布減少,E-cadherin的表達增加。然后,我們研究了MIR210HG是否通過改變miR-337-3p/137-HMGA2軸來影響EMT進展。Western blotting和免疫熒光結(jié)果顯示,MIR210HG的下調(diào)降低了Ishikawa和HEC1A細胞中HMGA2蛋白的表達,而加入miR-337-3p/137抑制劑后,對HMGA2、b-catenin和E-cadherin表達的抑制作用被逆轉(zhuǎn)(圖6G和6H)。

7) MIR210HG通過miR-337-3p和miR-137促進腫瘤進展
為了確定MIR210HG下調(diào)對Ishikawa和HEC-1A細胞系惡性行為的抑制作用是否通過miR-337-3p/137介導(dǎo),我們進行了功能挽救實驗。結(jié)果證實MIR210HG沉默對Ishikawa和HEC-1A細胞遷移和侵襲的惡性抑制作用被miR-337-3p/137敲低顯著逆轉(zhuǎn)(圖7A-7E)。這表明MIR210HG通過miR-337-3p/137調(diào)控Ishikawa和HEC-1A細胞的惡性生物學(xué)行為。

8)抑制MIR210HG和miR-337-3p/137過表達抑制腫瘤生長
我們測定了MIR210HG和miR-337-3p/137在體內(nèi)腫瘤模型中的功能。裸鼠實驗中,每組實驗小鼠3只。結(jié)果表明與對照組相比,轉(zhuǎn)染sh-MIR210HG并過表達miR-337-3p和miR-137降低了腫瘤體積(圖8A)。轉(zhuǎn)染shMIR210HG并同時過表達miR-337-3p和miR-137的組腫瘤體積最小。免疫組化分析顯示,與單獨轉(zhuǎn)染agomir-337-3p/137和sh-MIR210HG組相比,共轉(zhuǎn)染組HMGA2和Ki-67的表達較低(圖8B)。
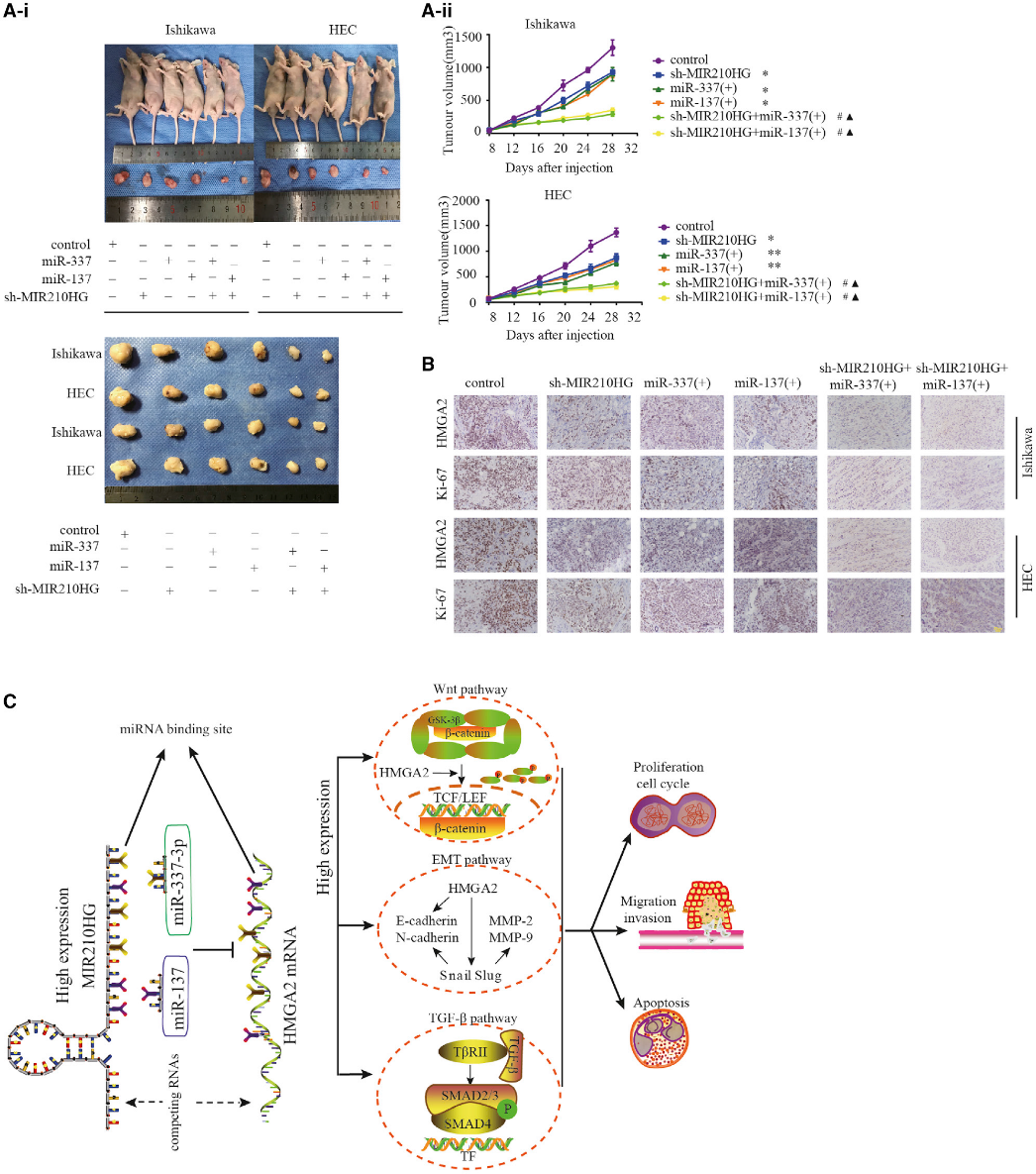
結(jié)論:MIR210HG在子宮內(nèi)膜癌患者中是一個獨立的預(yù)后因素,干擾MIR210HG的體內(nèi)外表達可抑制子宮內(nèi)膜癌細胞的惡性表型。MIR210HG-miR-337-3p/137被發(fā)現(xiàn)介導(dǎo)HMGA2調(diào)節(jié)子宮內(nèi)膜癌細胞惡性行為的能力。MIR210HG-miR-337-3p/137-HMGA2軸是新的子宮內(nèi)膜癌分子預(yù)后標志物和潛在的治療靶點。