lncRNA VS骨形成
導語:骨是一個動態(tài)器官,通過破骨細胞和成骨細胞的協(xié)調產生自我修復潛力。衰老過程中骨的自我修復能力明顯受損。間充質骨祖細胞(OPCs)向骨形成表面的遷移是成骨細胞生成的初始步驟,OPCs的遷移能力在衰老過程中是否受到損害,以及它如何促成衰老相關的骨形成減少仍不清楚。今天,lncRNA粉墨登場,演繹著不一樣的精彩故事。
參考文獻:Targeting long noncoding RNA PMIF facilitates osteoprogenitor cells migrating to bone formation surface to promote bone formation during aging (IF=8.579)
技術路線:

結果
1. 衰老過程中骨形成減少伴隨OPCs向骨形成表面遷移減少,lnc-PMIF表達升高
收集衰老小鼠模型的骨標本,分析動態(tài)骨組織形態(tài),發(fā)現(xiàn)老年小鼠的礦化沉積率(MAR)和骨形成率(BFR/BS)均顯著降低,表明衰老期間小鼠的骨形成減少。從小鼠中分離出BMSCs,RNA-Seq分析發(fā)現(xiàn)老年BMSCs中遷移相關基因均下調。老年和年輕小鼠BMSCs成骨誘導7d后ALP活性和成骨誘導14d后鈣礦物質沉積相當,表明BMSCs的成骨潛能在衰老過程中沒有改變。再次從老年和年輕小鼠中分離BMSCs,Dil染色劑標記后體外增殖,脛骨內注射到另一批年輕小鼠中,三天后收獲脛骨對成骨標志物Runx2進行熒光免疫染色。發(fā)現(xiàn)老年BMSCs處理的小鼠中Dil標記細胞降低,表明老化過程中OPCs向骨形成表面的遷移能力降低。大部分遷移到骨形成表面的Dil標記細胞表達Runx2,表明OPCs遷移到骨形成表面發(fā)生成骨分化,有助于體內骨形成。與年輕BMSCs相比,老年BMSCs的體外遷移能力明顯受損,Macf1顯著下調,從Macf1基因位點轉錄而來的長鏈非編碼RNA PMIF(即lnc-PMIF)表達顯著增高。上述提示,衰老過程中小鼠骨形成的減少伴隨著OPCs向骨形成表面遷移的減少和OPCs中lnc-PMIF表達的升高。

2. Lnc-PMIF抑制OPCs遷移到骨形成表面
在成骨細胞分化過程中,lnc-PMIF在早期增加,在礦化啟動后減少,表明lnc-PMIF可能在成骨分化的早期階段發(fā)揮作用。過表達/敲低lnc-PMIF不影響OPCs的增殖和成骨分化。敲低lnc-PMIF的細胞遷移減弱,敲低組細胞處理的小鼠脛骨中Dil標記細胞數(shù)量升高,小鼠中lnc-PMIF表達更高。這些表明lnc-PMIF可抑制OPCs向骨形成表面的遷移,而不干擾其增殖和成骨潛能。

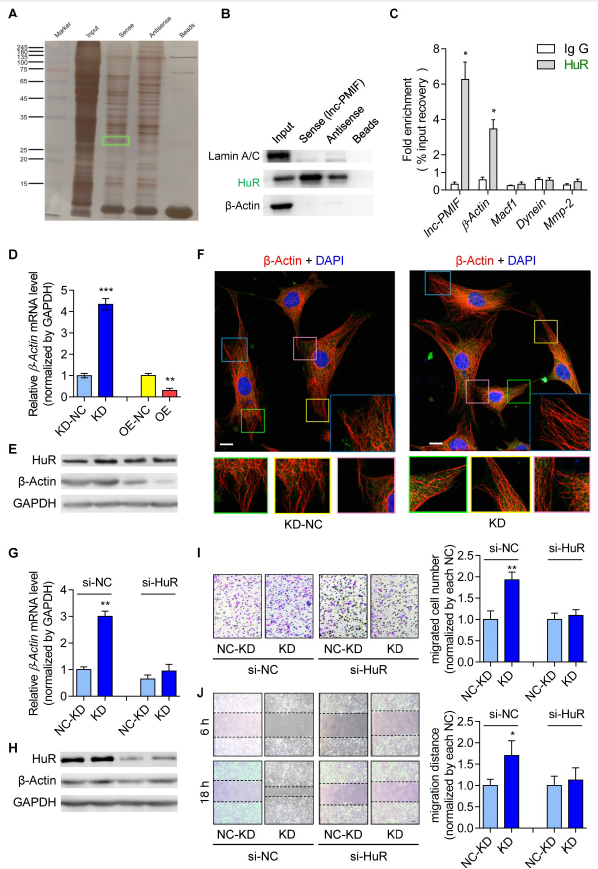
3.Lnc-PMIF與HuR相互作用抑制β-actin的表達以抑制OPC遷移
為探索Lnc-PMIF介導的OPC遷移的調節(jié)機制。RNA pull down-MS實驗尋找lnc-PMIF的相互作用蛋白,鑒定出一種RNA結合蛋白HuR,WB證實HuR在生物素標記的lnc-PMIF細胞裂解的下拉組分中富集,RNA免疫沉淀(RIP)試驗lnc-PMIF與HuR的結合。還檢測到β-肌動蛋白顯著升高,與免疫沉淀的HuR蛋白結合。接下來研究HuR-β-actin介導的調節(jié)機制是否參與OPC的遷移調節(jié)。β-actin的mRNA和蛋白表達以及細胞遷移能力在HuR基因過表達的細胞中顯著增加, HuR基因敲低細胞中降低。β-肌動蛋白的mRNA和蛋白表達在lnc-PMIF過表達細胞中顯著下調,但在lnc-PMIF基因敲除細胞中顯著上調,表明lnc-PMIF能抑制OPCs中β-actin的表達,基因過表達/敲除lnc-PMIF不改變HuR的蛋白表達。在前述lnc-PMIF敲低細胞中敲低HuR基因,細胞中β-actin的mRNA和蛋白表達顯著下調,細胞遷移能力也受到損害,這些數(shù)據(jù)表明lnc-PMIF可與HuR相互作用,抑制β-actin的表達來抑制OPC遷移。
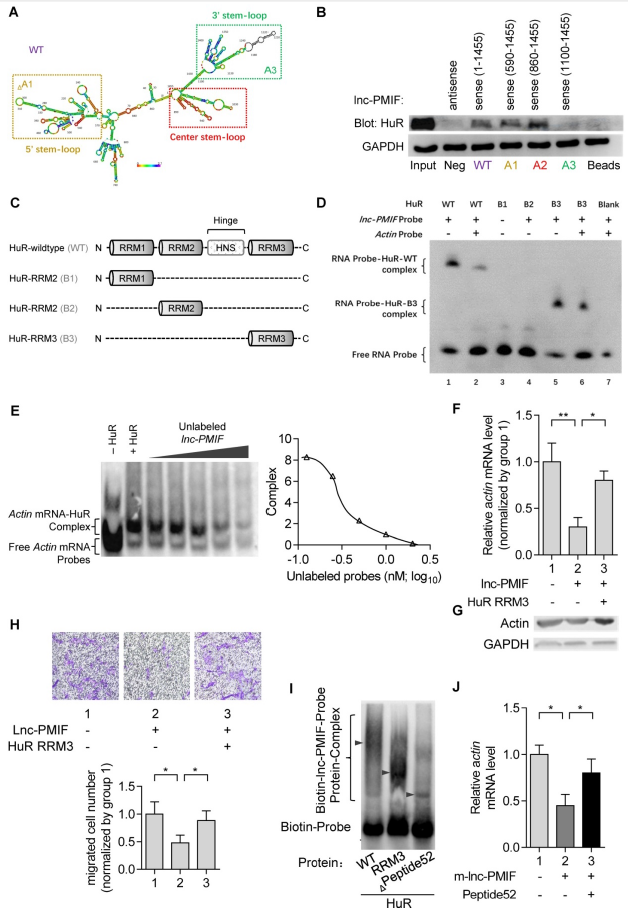
4. lnc-PMIF與HuR的RRM3結合,中斷HuR-β-actin mRNA相互作用,抑制β-actin表達,抑制OPC遷移
為闡明lnc-PMIF如何與HuR相互作用調節(jié)β-actin表達和OPC遷移,首先通過生物信息學分析預測了lnc-PMIF的二級結構,并合成生物素化的全長lnc-PMIF(WT)和截短的lnc-PMIF突變體,分別為突變體A1(無5’莖環(huán)的正義)、突變體A2(有中心莖環(huán)和3’莖環(huán)的正義)和突變體A3(僅有3’莖環(huán)的正義1100-1455 bp),轉染MC3T3-E1細胞,并進行標記RNA鏈霉親和素下拉試驗。蛋白免疫印跡分析顯示,HuR存在于轉染WT lnc-PMIF、截短lnc-PMIF突變體A1或截短lnc-PMIF突變體A2的細胞的下拉部分中,但在轉染截短lnc-PMIF突變體A3的細胞的下拉部分中不存在。這些數(shù)據(jù)表明,lnc-PMIF的中心莖環(huán)足以實現(xiàn)lnc-PMIF和HuR之間的相互作用。
接下來,我們合成了野生型HuR蛋白(WT)和HuR蛋白的截短突變體(即分別為單獨HuR的RNA識別基序(RRM)1的突變體B1、單獨HuR RRM2的突變體B2和單獨HuR RRM3的突變體B3)(圖4 C)。同時,我們設計并合成了生物素標記的或未標記的探針,分別特異性靶向lnc-PMIF的中心-莖環(huán)和生物素標記的或未標記的探針,特異性靶向之前報道的與HuR蛋白相互作用的β-actin mRNA的序列。RNA電泳遷移率變動分析(EMSA)發(fā)現(xiàn)只有當生物素標記的lnc-PMIF探針與WT HuR或截短突變體B3共孵育時,才能檢測到lnc-PMIF-HuR復合物,這表明HuR的RRM3可介導HuR和lnc-PMIF之間的相互作用。lnc-PMIF與β-actin mRNA競爭與HuR相互作用。過表達HuR-RRM3后細胞中β-actin的mRNA和蛋白表達下降顯著上調,遷移活性增強。所有這些表明lnc-PMIF可與HuR的RRM3結合,中斷HuR-β-actin的相互作用,抑制β-actin的表達,抑制OPC的遷移。
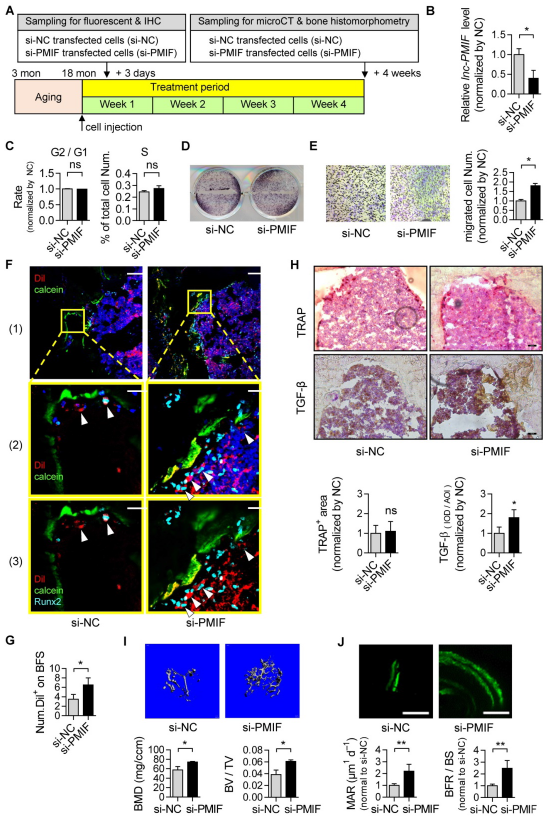
5. 沉默lnc-PMIF促進老年OPCs向骨形成表面遷移,促進老年小鼠骨形成
BMSCs中沉默lnc-PMIF,增殖無變化,OPCs成骨能力不改變,細胞遷移數(shù)量高,表明沉默lnc-PMIF可增強老年OPCs的遷移能力,而不干擾老年OPCs的體外增殖和成骨分化。接受si-PMIF處理的老年BMSCs的小鼠中骨形成表面Dil +細胞數(shù)顯著升高,提示沉默lnc-PMIF可促進老年OPCs在體內向骨形成表面遷移,大部分遷移的Dil +細胞中檢測到Runx2表達,表明si-PMIF處理的老年BMSCs遷移到骨形成表面發(fā)生正常的成骨分化,這將有助于小鼠的骨形成。接受si-PMIF處理的老年BMSCs小鼠骨表面TGF-β+細胞的平均積分光密度顯著更高,而TRAP+面積無顯著差異,表明si-PMIF處理的老化BMSCs可能影響細胞募集,而不是骨吸收。上述si-PMIF或si-NC處理的老年BMSCs對老年雄性小鼠進行脛骨內注射,4周后接受si-PMIF處理的老年BMSCs的小鼠骨量更高,脛骨干骺端微結構更好,BMD和BV/TV更高,MAR和BFR/BS更高。這些結果證明,老年BMSCs敲低lnc-PMIF可促進骨形成。
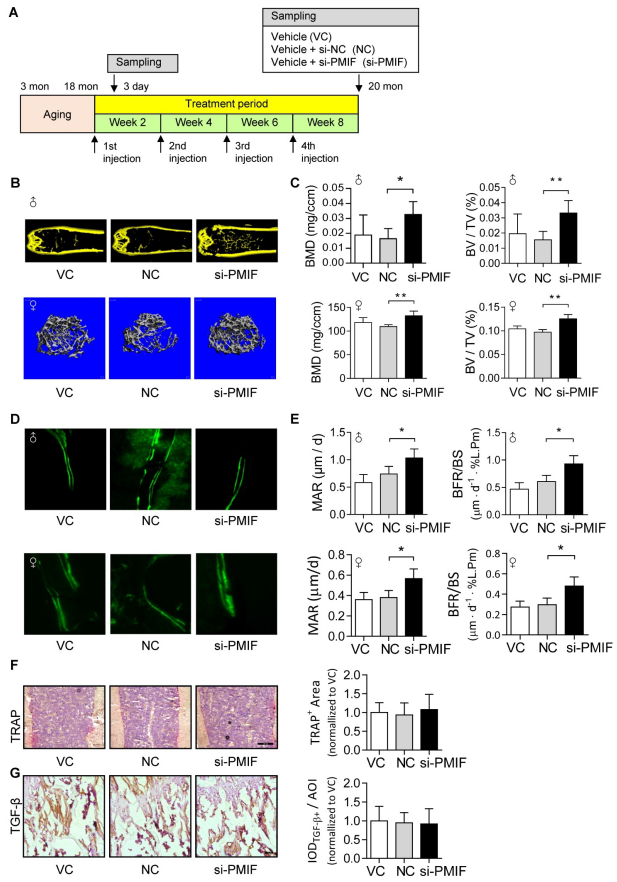
6. 靶向遞送lnc-PMIF siRNA接近骨形成表面的OPCs促進老年小鼠骨形成
接下來評價lnc-PMIF在骨形成表面周圍的老年OPCs中系統(tǒng)靶向沉默對衰老相關骨質疏松的影響。為促進lnc-PMIF siRNA(即si-PMIF)接近骨形成表面周圍的OPCs,si-PMIF或si-NC被包裹在骨形成表面靶向遞送系統(tǒng)(即,(DSS6)-脂質體)。21月齡自然衰老的雄性和雌性小鼠靜脈注射 (DSS6)-脂質體-si-PMIF、(DSS6)-脂質體-si-NC或(DSS6)-脂質體單藥(溶劑)。si-PMIF處理組Gli1 +細胞中lnc-PMIF的表達顯著降低,si-PMIF處理組兩種性別小鼠的骨量更高,脛骨干骺端微結構更好,BMD和BV/TV更高,MAR和BFR/BS較高。這些發(fā)現(xiàn)表明,靶向沉默骨形成表面周圍OPCs中的lnc-PMIF可以促進衰老相關骨質疏松小鼠的骨形成。