






瘧疾?不可怕,先來看看m6A甲基化演個什么角
參考文獻(xiàn):
Baumgarten Sebastian., Bryant Jessica M., Sinha Ameya., Reyser Thibaud., Preiser Peter R., Dedon Peter C., Scherf Artur., (2019). Transcriptome-wide dynamics of extensive mA mRNA methylation during Plasmodium falciparum blood-stage development. Nat Microbiol, 08. IF:14.3
導(dǎo)語:
瘧疾奪去了許多人的生病,它的發(fā)病機(jī)理是由于惡性瘧原蟲在人體紅細(xì)胞內(nèi)的無性繁殖造成的,這個過程發(fā)生在瘧疾蟲短暫的48小時生命中,并需要精確的基因表達(dá)級聯(lián)才能完成。雖然目前已經(jīng)在轉(zhuǎn)錄后水平觀察到這種反應(yīng),但是在轉(zhuǎn)錄組范圍的介導(dǎo)機(jī)制并不清楚。因此作者在惡性瘧原蟲轉(zhuǎn)錄組中鑒定了mRNA修飾,并在血液階段發(fā)育過程中對N6-甲基腺苷(m6A)進(jìn)行了全面表征。該研究展示了瘧疾寄生蟲中廣泛的m6A mRNA甲基化程序的獨(dú)特特征,并揭示其在動態(tài)微調(diào)單細(xì)胞真核生物的轉(zhuǎn)錄級聯(lián)中的關(guān)鍵作用。
結(jié)果分析:
1、鑒定惡性瘧原蟲在blood-stage階段的動態(tài)m6A甲基化程序
圖示惡性瘧原蟲在人紅細(xì)胞內(nèi)的無性繁殖周期,包括紅細(xì)胞侵入、宿主細(xì)胞重構(gòu)、分裂復(fù)制和紅細(xì)胞排出(圖1a)。在分析紅細(xì)胞內(nèi)的發(fā)展周期(IDC)中鑒定出10個在mRNA的修飾(圖1b,c,d)。雖然m6A甲基化的程度總體都很高,但是在瘧原蟲中更高(圖1e)。
惡性瘧原蟲基因組編碼m6A mRNA甲基轉(zhuǎn)移酶的高度保守(圖2a),這個保守去編碼的蛋白命名為PfMT-A70,構(gòu)建了包含HA標(biāo)簽的PfMT-A70蛋白表達(dá)細(xì)胞系(圖1b)。對PfMT-A70-HA進(jìn)行免疫沉淀,然后進(jìn)行LC-MS,鑒定出16種特異性蛋白(圖2c,d)。三種惡性瘧原蟲的轉(zhuǎn)錄水平都在12和24h.p.i時達(dá)到高峰,比m6A/A比率最高時早一點(diǎn)(圖1e,2e)。
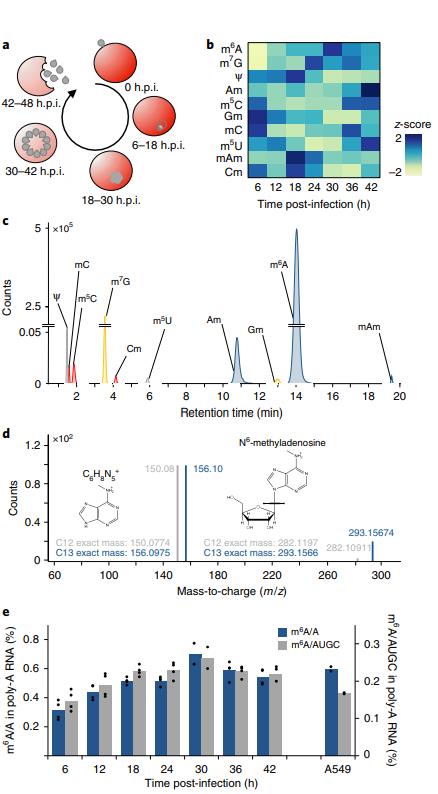
圖1惡性瘧原蟲紅細(xì)胞內(nèi)部(IDC)發(fā)mRNA修飾全局動態(tài)
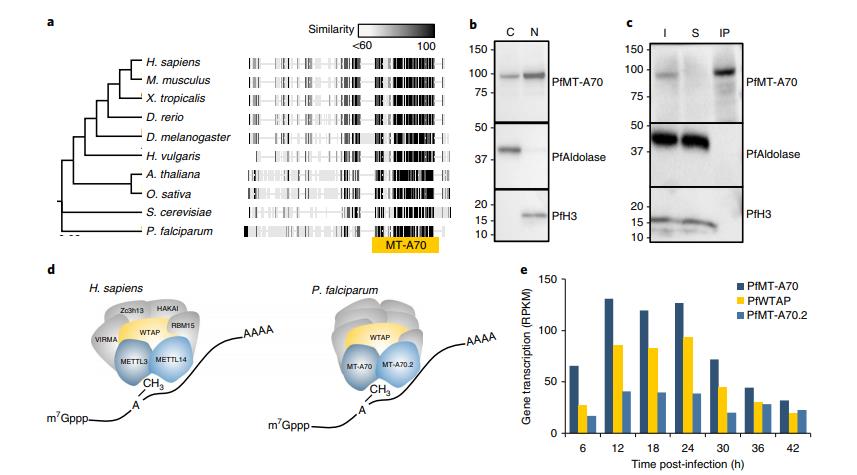
圖2 惡性瘧原蟲m6A書寫復(fù)合體的特征
2、PfMT-A70的CRISPR干擾降低了整體m6A mRNA甲基化
對于PfMT-A70的CRISPRi,我們將其啟動子定位于轉(zhuǎn)錄起始位點(diǎn)下游和翻譯起始位點(diǎn)上游~100bp,gRNA與非模板鏈互補(bǔ),非特異性的gRNA表達(dá)細(xì)胞系作為陰性對照(圖3a)。gPfMT-A70靶向dCas9染色質(zhì)免疫沉淀,然后測序(ChIPseq)在靶點(diǎn)顯示出強(qiáng)大且高度特異性的富集,其與富含H3K9ac5和H2A.Z33的核小體缺失區(qū)域(FAIRE)重疊(即標(biāo)志惡性瘧原蟲啟動子區(qū)域;圖3b,c)。RT-qPCR表明整個IDC中PfMT-A70轉(zhuǎn)錄水平顯著下調(diào),在12h.p.i時敲低水平為~15%,在24和36h.p.i時達(dá)到40% (圖3d,頂部)。 雖然PfMT-A70 CRISPRi對寄生蟲生長沒有明顯的影響,但LC-MS / MS顯示在所有三個時間點(diǎn)m6A / A整體水平顯著下降10-30%(圖3d, 底部),進(jìn)一步確定PfMT-A70是惡性瘧原蟲m6A書寫復(fù)合體的組成部分。
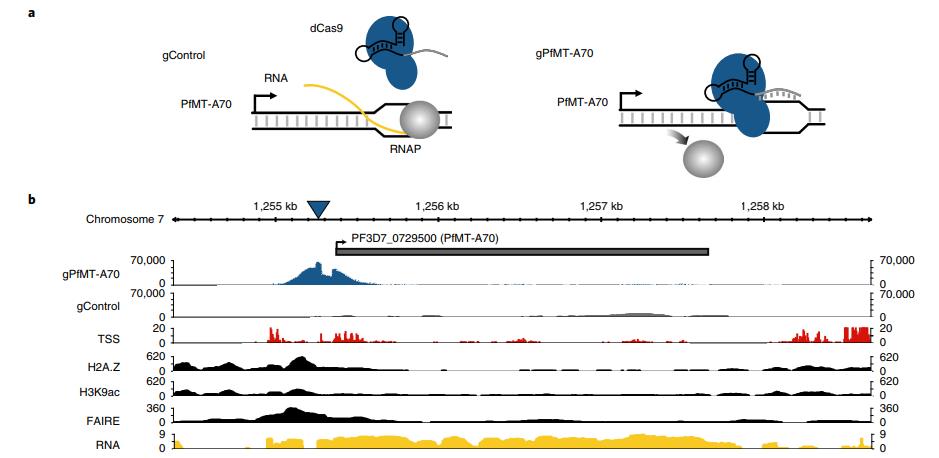
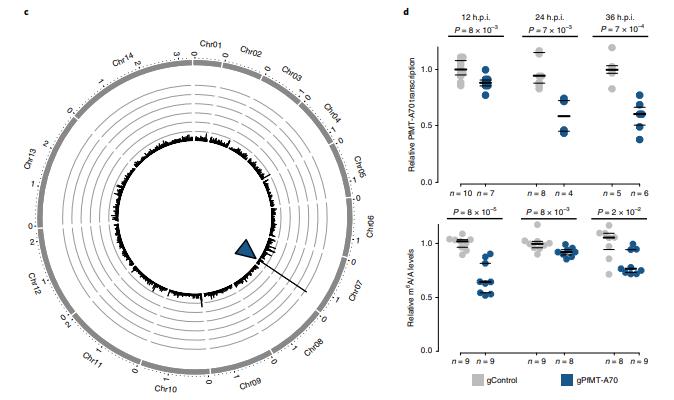
圖3通過CRISPRi敲低PfMT-A70 m6A甲基轉(zhuǎn)移酶
3、在IDC期間,mRNA轉(zhuǎn)錄物差異地m6A-甲基化
作者使用m6A-甲基化mRNA免疫沉淀(IP)和m6A-seq測定了IDC的轉(zhuǎn)錄物特異性水平上m6A mRNA甲基化的動態(tài)。 m6A-IP及LC-MS / MS顯示抗體特異性高(圖4a)。與后生動物和植物中m6A位點(diǎn)的拓?fù)浣Y(jié)構(gòu)相反,惡性瘧原蟲中的m6A峰不會在終止密碼子和3'UTR附近富集,而表現(xiàn)出向5'末端定位的趨勢(圖4b,c)。作者搜索了與m6A峰相關(guān)的序列,發(fā)現(xiàn)GGACA基序最顯著富集,該基序類似于在酵母和人m6A位點(diǎn)處鑒定的RGAC和DRACH(D = G / A / U,R = G / A,H = C / A / U)基序(圖4d)。GAC基序集中在m6A峰頂周圍的±20nt區(qū)域(圖4e),表明特定m6A甲基化的某些背景依賴性。
在整個IDC中,具有至少一個m6A峰的轉(zhuǎn)錄基因總數(shù)略有增加(圖4f)。 然而,m6A富集(即m6A-IP / m6A-輸入),或來自含有特定m6A峰的一個基因的轉(zhuǎn)錄物的部分,在整個IDC中發(fā)生了廣泛的變化(圖4g)。 大多數(shù)m6A峰僅在一個時間點(diǎn)達(dá)到最大m6A富集,表明轉(zhuǎn)錄特異性甲基化確實是一種主動調(diào)節(jié)機(jī)制。
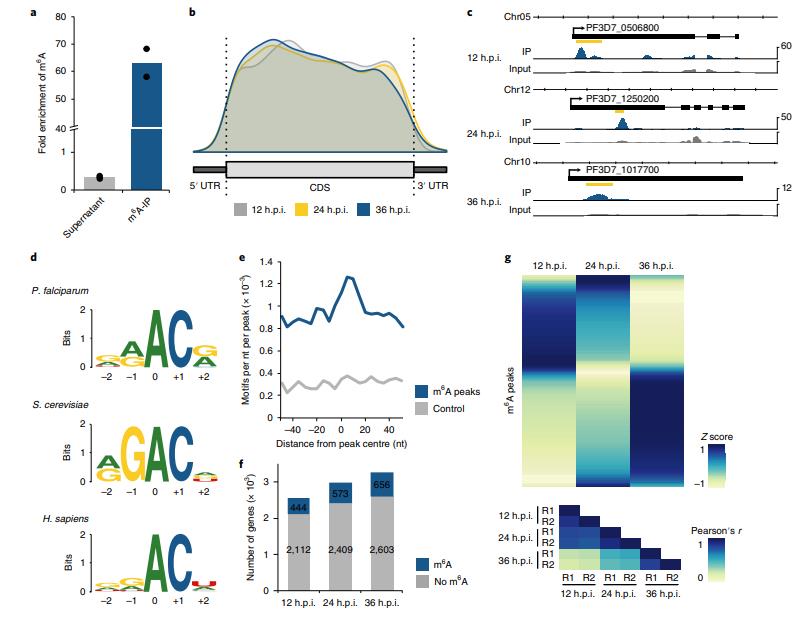
圖4 惡性瘧原蟲IDC期間的差異m6A甲基化
4、m6A與mRNA穩(wěn)定性和翻譯效率成反比關(guān)系
為了確定m6A對個體轉(zhuǎn)錄本的影響,首先比較gControl和gPfMT-A70寄生蟲之間各個m6A峰的m6A富集,并發(fā)現(xiàn)gPfMT-A70細(xì)胞系中所有時間點(diǎn)的富集都顯著減少(圖5a,b,頂部)。通過LC-MS / MS測量的gPfMTA70寄生蟲中全局m6A / A水平的降低來反映。(圖3d,底部)。 gControl和gPfMT-A70在每個時間點(diǎn)顯示高度相關(guān)性(圖5b,底部)。
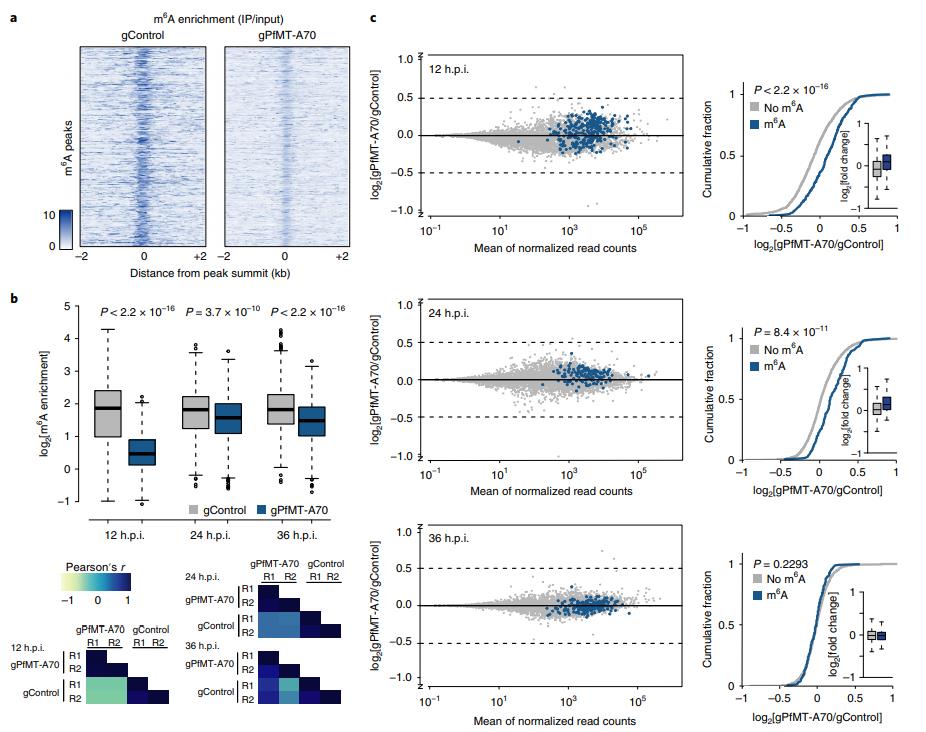
圖5 PfMT-A70基因敲除導(dǎo)致m6A甲基化轉(zhuǎn)錄本的上調(diào)
m6A-甲基化轉(zhuǎn)錄物具有比非甲基化轉(zhuǎn)錄物顯著更低的mRNA穩(wěn)定性(圖6a,左)。 這些差異在12和24h.p.i顯著,但在36h.p.i時不顯著。在12和24h.p.i時,‘m6A敏感’轉(zhuǎn)錄物的mRNA穩(wěn)定性甚至更低(m6A富集減少兩倍,轉(zhuǎn)錄本豐度增加(圖6a,右)。比較整個IDC中m6A-甲基化和非甲基化轉(zhuǎn)錄物之間的翻譯效率(TE)。在每個發(fā)育階段,m6A甲基化與轉(zhuǎn)錄物的翻譯效率顯著降低相關(guān)(圖6b,左)。這種相關(guān)性在12和24h.p.i時最為明顯,但在36h.p.i時仍然顯著。 與mRNA穩(wěn)定性所見的模式類似,在12和24h.p.i時m6A甲基化和‘m6A敏感’轉(zhuǎn)錄物子集之間的翻譯效率的顯著降低(圖6b,右)。
5、惡性瘧原蟲編碼進(jìn)化上保守的,多樣化的m6A結(jié)合蛋白
惡性瘧原蟲基因組編碼含有YTH結(jié)構(gòu)域的兩種不同蛋白質(zhì):PfYTH.1(PF3D7_1419900,此前注釋為‘切割和多聚腺苷酸化特異性因子亞基4’,CPSF4)和PfYTH.2(PF3D7_0309800,圖6c)。PfYTH.2與其他已知m6A讀取蛋白的沒有總體相似性。兩種蛋白質(zhì)在IDC早期高度表達(dá)(圖6d),并且PfYTH.1的異位表達(dá)揭示了蛋白質(zhì)在細(xì)胞核和細(xì)胞質(zhì)中的定位(圖6e)。與IDC期間的動態(tài)表達(dá)相比,PfYTH.2在寄生蟲的傳播階段表現(xiàn)出甚至更高的表達(dá),即配子體和唾液腺子孢子,后者也高度表達(dá)PfMT-A70(圖6d)。
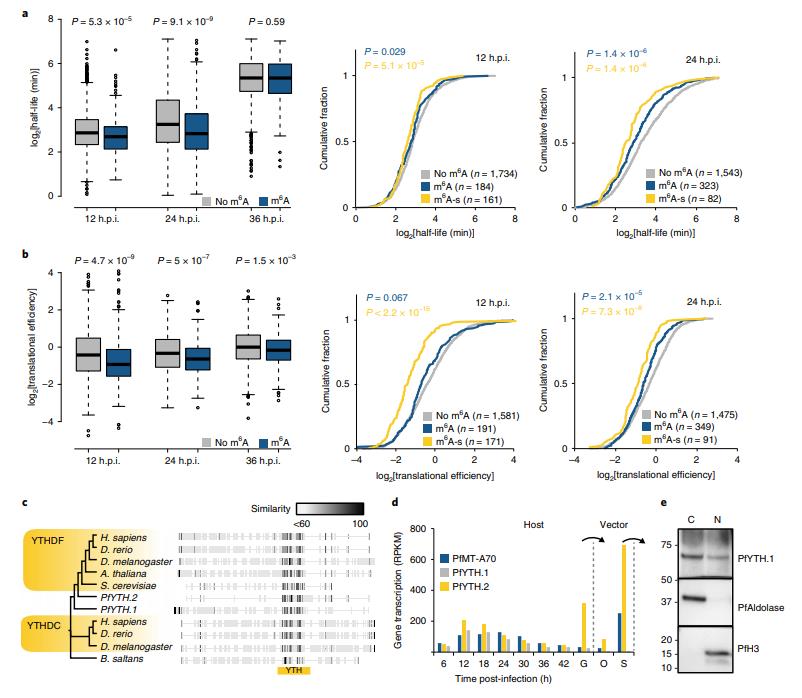
圖6 m6A與mRNA穩(wěn)定性及翻譯效率的關(guān)系
結(jié)語:
該研究揭示了惡性瘧原蟲中基因表達(dá)的另一層動態(tài)和廣泛的轉(zhuǎn)錄后調(diào)節(jié)。 m6A mRNA甲基化的核心特征的保守性使惡性瘧原蟲成為研究m6A甲基化和基因轉(zhuǎn)錄之間相互作用的優(yōu)良系統(tǒng)。此外,該研究將m6A作為瘧疾寄生蟲“上皮轉(zhuǎn)錄組”代碼的主要參與者并為瘧疾寄生蟲的藥物的發(fā)展開辟了新的途徑。